

Descargar este artículo en formato PDF
Götz, Christopher M.
2005 El consumo de vertebrados en tres grupos habitacionales de Siho, Yucatán. En XVIII Simposio de Investigaciones Arqueológicas en Guatemala, 2004 (editado por J.P. Laporte, B. Arroyo y H. Mejía), pp.781-797. Museo Nacional de Arqueología y Etnología, Guatemala.
76
EL CONSUMO DE VERTEBRADOS EN TRES GRUPOS
HABITACIONALES DE SIHO, YUCATÁN
Christopher M. Götz
Palabras clave:
Arqueología Maya, México, Yucatán, Siho, análisis de fauna, arqueozoología, contenido de basureros, paleo-nutrición
Siho está ubicado en la parte noroeste del estado de Yucatán, México, aproximadamente 30 km al oeste de la Costa del Golfo. Ha sido investigado desde el 2001 por el Proyecto Arqueológico Siho, de la Facultad de Ciencias Antropológicas de la Universidad Autónoma de Yucatán (Cobos et al. 2002). El asentamiento prehispánico estuvo ocupado desde el Preclásico Medio hasta el Clásico Terminal (600 AC – 1100 DC), con una aparente interrupción de ocupación durante el Preclásico Tardío y un apogeo poblacional durante el Clásico Tardío. Durante el Clásico Terminal (800-1100 DC), se agregaron y modificaron algunas estructuras, mientras que la ocupación disminuyó considerablemente a partir de esta época (Cobos et al. 2002:92-93).
El presente trabajo mostrará los resultados del análisis zoo-arqueológico de elementos esqueléticos de animales vertebrados recuperados entre 2001 y 2003 en tres basureros prehispánicos del sitio. El trabajo investiga el aprovechamiento alimenticio de animales vertebrados por los habitantes prehispánicos del sitio. Los tres contextos zoo-arqueológicos analizados en este trabajo se asocian a las estructuras habitacionales 5D2, 5D16 y 5D13/5D14 respectivamente, todas atribuidas al nivel socio-económico de élite (Figura 1). El análisis zoo-arqueológico compara los tres basureros entre sí y se indagan los patrones alimenticios de la élite del sitio entre el Clásico Tardío y el Clásico Terminal al contextualizar los resultados zoo-arqueológicos con la información arqueológica del sitio. Los resultados se contrastan tanto a nivel micro regional como macro regional, con análisis zoo-arqueológicos de sitios como Dzibilchaltun y Yula en las Tierras Bajas del Norte (Wing 1980; Carr 1995), así como Ceibal y el área de Petexbatun en las Tierras Bajas Centrales (Pohl 1976, 1990; Emery 1997), para evidenciar diferencias y similitudes en cuanto a esta práctica cultural. De esta manera se presentará un análisis acerca de la explotación del medio ambiente faunístico y de las estrategias de obtención de alimento en las Tierras Bajas Mayas del Norte durante el periodo Clásico de acuerdo a los conocimientos actuales.
METODOLOGÍA DE CAMPO Y DE LABORATORIO
Los materiales faunísticos analizados de Siho proceden de los pozos de prueba y de las excavaciones horizontales. La tierra matriz de todas las excavaciones fue cernida completamente con un cernidor de aperturas de 5 mm, por lo que se considera que la representatividad de presencia o ausencia de pequeños huesos y fragmentos óseos debe considerarse igual para las tres áreas de comparación.
La identificación taxonómica y osteológica de los materiales de Siho se efectuó a través de la comparación de los especímenes arqueológicos con otros de una colección moderna comparativa, con una base fotográfica digital de especímenes esqueléticos modernos de la región Maya (EA.FLMNH 2003), y por medio de manuales de osteología faunística para mamíferos, aves y reptiles (Olsen 1968; Gilbert et al. 1980, 1985). La terminología taxonómica se basa en recientes publicaciones sobre la fauna regional (Reid 1997; Lee 2000; Peterson et al. 1973).
Junto con la identificación taxonómica se denominó el elemento esquelético representado y el lado al que corresponde un elemento par. De esta manera se calculó el número de especímenes identificados por taxon (NISP, por sus siglas en inglés), el número mínimo de individuos (MNI, por sus siglas en inglés), y la distribución de fragmentos por porción corporal de cada taxon en los contextos al respecto (frecuencia esquelética).
Una limitante del análisis de los elementos esqueléticos faunísticos de Siho es la reducida cantidad de fragmentos óseos encontrados. En total se recuperaron 361 fragmentos óseos de animales vertebrados en todo el sitio. Para los contextos discutidos aquí se considerarán 160 fragmentos identificados a nivel de género o especie, además de 28 fragmentos identificados a nivel de familia y orden, resultando en un número de 32 individuos mínimos (Figura 2). Las comparaciones de los contextos se basan por esta razón en el número de especímenes identificados por taxon (NISP; Gautier 1984), ya que el número mínimo de individuos para la mayoría de los taxa es demasiado bajo para poder mostrar diferencias o semejanzas significativas. Gracias a que la fragmentación ósea de los taxa semejantes siguió un patrón similar en los contextos comparados directamente, se considera que las cuantificaciones del NISP son válidas entre otros (O’Connor 2000:56), y pueden, junto con las representaciones de la frecuencia esquelética, expresar las diferencias encontradas en los tres contextos faunísticos.
En cuanto a las interpretaciones posteriores hay que tomar en cuenta, desde luego, que la el NISP expresa únicamente los valores cuantitativos dados por los contextos mismos, sin que se puedan hacer aún estimaciones acerca de parámetros poblacionales de la fauna prehispánica del sitio. El contexto arqueológico de los datos faunísticos se basa en los informes del Proyecto Arqueológico Siho (Cobos et al. 2002; Cobos et al. 2004), así como en los análisis cerámicos (Cobos et al. 2004; Jiménez 2004).
EL CONJUNTO DE LA ESTRUCTURA 5D2
La Estructura 5D2 encierra desde el oeste, la plaza central del sitio formada además por las Estructuras 5D1 (este), 5D7 (sur) y 5D17 (norte). Durante los trabajos del Proyecto Arqueológico Siho se excavaron únicamente las Estructuras 5D2 y 5D7. La Estructura 5D1 fue probablemente la pirámide principal del sitio (Maler 1997:240). La Estructura 5D17 fue al parecer una estructura habitacional de élite de tipo palacio (Cobos et al. 2002), mientras que la Estructura 5D7 se caracteriza como edificio auxiliar con funciones habitacionales (Tun 2004). La Estructura 5D2 es un edificio de tipo palacio de dos cuartos abovedados (Dunning 1993:1). Se fecha arquitectónicamente al estilo Proto-Puuc entre 550 y 710 DC (Cobos et al. 2002:15; Fernández y Peniche, en Cobos et al. 2004:45), una fecha reforzada por el hallazgo de un glifo indicando el año 652 DC (Lacadena, en Cobos et al. 2004). Al parecer, la estructura, de carácter habitacional de élite, estuvo en uso durante entre el Clásico Tardío y el Clásico Terminal.
Los materiales faunísticos procedentes de la Estructura 5D2 fueron encontrados en un basurero ubicado a lo largo del muro norte del basamento, acumulado según los análisis cerámicos entre las facetas tempranas del Clásico Tardío y el Clásico Terminal (Figura 1; Fernández y Peniche, en Cobos et al. 2004:50). Los materiales procedentes del Clásico Tardío (550-800 DC), en la mayoría de los lotes procedentes de la capa II, se asocian arquitectónicamente a la construcción del edificio, mientras que los materiales del Clásico Terminal (800-1050/1100 DC), de la capa l y superficie en la mayoría de los lotes, se relacionan a remodelaciones arquitectónicas del edificio (Jiménez 2004). En total, se encontraron 100 fragmentos óseos de animales vertebrados en el basurero de la Estructura 5D2, 93 de éstos atribuibles al Clásico Tardío y siete al Clásico Terminal.
Los fragmentos identificados del Clásico Tardío abarcan, en diferentes proporciones, a una especie de ave grande no identificada con exactitud, posiblemente hocofaisán (Crax rubra), o pavo (Meleagris ocellata); peces marinos como bagre marino (Arius felis) y manta raya (Rayiformes); reptiles terrestres como serpientes cascabel (Crotalus durissus), y boa (Boa constrictor); iguana negra (Ctenosaura similis); y tortuga (Testudinae cf. Terrapene); así como mamíferos como venados temazate (Mazama americana), y cola blanca (Odocoileus virginianus; Figuras 2 y 3). Todos estos animales se encuentran actualmente en la región de Siho y se consideran endémicos (Reid 1997; Lee 2000; Peterson 1973). En las capas del Clásico Terminal se pudieron identificar únicamente fragmentos de venado sin poder precisar especie o género debido a una fuerte erosión (Cervidae n.d.), así como de venado cola blanca (Odocoileus virginianus; Figura 2).
La mayoría, tanto en los fragmentos del Clásico Tardío como del Clásico Terminal, se evidenció una gran cantidad de huellas de fuego y rupturas en hueso fresco. El venado cola blanca y la iguana negra están representados por todo el esqueleto (cabeza, esqueleto axial, extremidades y patas), mientras que los demás animales únicamente aparecen por partes seleccionadas del cuerpo, ya sea por las extremidades (ave), el esqueleto axial (tortuga terrestre, serpientes, manta raya), o la cabeza (bagre; Figura 3). El bagre marino se presenta en forma de unos pocos otolitos (o estatocistos), y una espina dorsal, indicando que hubo al menos algunas cabezas de estos peces en uso.
EL CONJUNTO DE LA ESTRUCTURA 5D16
La Estructura 5D16 forma en conjunto con las Estructuras 5D19 y 5D20, un pequeño grupo habitacional aproximadamente 200 m al noroeste del centro de Siho (Cobos et al. 2002:19). Las tres estructuras del conjunto fueron excavadas por el Proyecto Arqueológico Siho. La Estructura 5D16 parece haber sido, debido a las dimensiones y su volumen de construcción, el edificio principal del grupo. El edificio está compuesto por bóvedas similares al estilo arquitectónico Oxkintok Temprano (300-550 DC), o Proto Puuc (550-710 DC), mientras que el basamento presenta rasgos del estilo Megalítico (300-600 DC; Fernández et al. 2003). Siguiendo las interpretaciones de Fernández et al. (2003), la Estructura 5D16 parece haber sido residencia de individuos de alto estatus durante el Clásico Tardío y Clásico Terminal. Las Estructuras 5D19 y 5D20 fueron construcciones de materiales perecederos sobre un fundamento de piedra, posiblemente destinados a labores de la preparación de alimentos y almacenaje de bienes.
A ambos costados de la escalinata de la Estructura 5D16 se encontraron acumulaciones de materiales interpretables como basureros (Figura 1). Estas dos acumulaciones de basura son, al parecer, contemporáneas y fechan al Clásico Terminal (Lilia Fernández Souza, comunicación personal 2004). Durante las excavaciones de la Estructura 5D16 no se encontró un basurero grande como en la Estructura 5D2 y las acumulaciones de la escalinata son la única manera de reconstruir el uso de fauna del conjunto mencionado. La cantidad de la basura y su ubicación hacen suponer que se trata de basureros preliminares o de barrido de la estructura, misma que se encontró prácticamente vacía al excavarse.
Los basureros preliminares o de barrido se caracterizan por ser acumulaciones relativamente pequeñas de basura cerca de las estructuras, en esquinas de plataformas o escalinatas (Chase y Chase 2000:69). Esto los diferencia del basurero de la Estructura 5D2 y del Pozo 5, mismos que reflejan basureros grandes. El basurero de barrido probablemente acumulaba la basura durante un tiempo, para luego tirarla en otro sitio, aún desconocido.
Se podría argumentar que los basureros de barrido contienen menos restos óseos de animales para evitar el olor de la descomposición de la carne adherida y que por esto son menos representativos en cuanto a la reconstrucción de costumbres alimenticias. Sin embargo, también el basurero de la Estructura 5D2, conteniendo una multitud de material óseo, estuvo ubicado cerca de la estructura, en donde se habrían desarrollado olores desagradables. Igualmente hay que considerar que los restos de preparación y consumo cárnico cerca de estructuras domésticas son generalmente cocidos, desarrollando menos olor en la descomposición. En este sentido, a pesar de la presunta diferencia en cuanto al tipo de basurero, se postula que ambos contextos sirven igualmente para la reconstrucción de los hábitos alimenticios de sus antiguos usuarios.
En total se encontraron 39 fragmentos óseos de animales vertebrados en los dos basureros, tratados a continuación como una sola unidad. Los restos proceden en su mayoría de animales pequeños, tales como roedores (Rodentia n.d. y Orthogeomys hispidus), iguanas negras (Ctenosaura similis) y serpientes (Boa constrictor); únicamente 15% de los restos proceden de perro (Canis familiaris), venado (Cervidae n.d.), y mamífero grande indeterminado (Figuras 2 y 4). Al igual que en el caso de la Estructura 5D2 se encontró una espina de bagre marino (Arius felis), que probablemente se utilizó como utensilio perforador. Los restos de roedor, tuza, serpiente y mamífero indeterminado muestran huellas de ruptura en fresco, además de contar, en algunos pocos casos, con huellas de fuego (roedor y mamífero indeterminado; Figura 4).
A pesar de la escasez de las huellas de fuego y la ausencia de las marcas de corte se interpreta el basurero aquí como resto de alimento de los habitantes de la estructura, argumentando que animales pequeños comúnmente se preparan usando todo el cuerpo (por ejemplo, pollos asados), permaneciendo los huesos cubiertos por piel y carne al cocinarse, por lo que quedarían menos huellas de fuego visibles. Las rupturas en fresco ocurrirían al partir el animal en porciones comestibles a través de la ruptura manual, sin que se tengan que utilizar herramientas cortantes.
EL BASURERO ENTRE LAS ESTRUCTURAS 5D13 Y 5D14
El Pozo de prueba No. 5 reveló una apertura circular en la plataforma entre las estructuras 5D13 y 5D14, que al parecer fue utilizada como basurero en tiempos prehispánicos (Figura 1). Las Estructuras 5D13 y 5D14 no se excavaron, pero su ubicación al este del centro, así como su forma y arquitectura sugiere una función habitacional relacionada a la élite (Rafael Cobos y Lilia Fernández Souza, comunicación personal 2004). La estratigrafía divide el basurero en cinco capas, fechándose las capas más profundas al Clásico Tardío (550-750/800 DC), y la capa I y el material de superficie al Clásico Terminal (Jiménez 2004).
En total, se recuperaron 155 fragmentos del Clásico Tardío y 42 fragmentos del Clásico Terminal. Las capas del Clásico Tardío revelaron restos de pavo del monte (Meleagris ocellata), perro (Canis familiaris), así como de venado cola blanca (Odocoileus virginianus), jabalí (Tayassuidae cf. Tayassu tajacu), de venado indeterminado (Cervidae n.d.), y de mamífero indeterminado (Figura 2). Similar al basurero de la Estructura 5D2 se presenta una amplia cantidad de rupturas en fresco, huellas de quemado y huellas de corte en los fragmentos óseos faunísticos, sobre todo en los restos de venado y mamífero indeterminado (Figuras 5 y 6).
INTERPRETACIÓN DE LAS COSTUMBRES ALIMENTICIAS EN SIHO DURANTE EL CLÁSICO
Los contextos faunísticos de las recientes excavaciones en Siho, Yucatán, revelan diferentes patrones y estrategias de la obtención de animales por la élite desde las facetas tempranas del Clásico Tardío hasta el Clásico Terminal. Los taxa identificados del Pozo 5 se limitan a animales grandes, básicamente procedentes de venado, jabalí, perro y pavo del monte. Los fragmentos de perro no muestran huellas de preparación, aunque antes se ha indicado haber encontrado únicamente huellas de corte y fuego en los metapodios, calcáneos y astrágalos de estos animales como prueba de su proceso alimenticio (Pohl 1990:164), elementos que están poco representados en nuestra muestra. En total, los animales del Pozo 5 son de mucha aportación cárnica, encontrándose en muchos contextos faunísticos de élite en sitios Mayas del Clásico. El venado, jabalí y pavo de monte representan aún hoy unas de las presas más preferidas de los cazadores rurales (Leopold 2000).
La mayoría de los fragmentos de venado procede de las extremidades traseras, seguidas por menos fragmentos del esqueleto axial y aún menos de las extremidades delanteras. El jabalí está principalmente representado por fragmentos de la cabeza. Estos patrones muestran una diferencia a las frecuencias esqueléticas de venados y jabalís descritos para las Tierras Bajas Centrales durante el Clásico Tardío. Tanto en Ceibal (Pohl 1976), como en la región de Petexbatun (Emery 1997:313), se asocian esencialmente las extremidades delanteras de venados, mientras que el esqueleto axial de venados se utilizaban por personas de rango menor (Pohl 1990:161-163), y la cabeza de jabalí parece asociarse a banquetes rituales.
Una alta incidencia de huellas de preparación en los huesos indica un aprovechamiento intensivo de los taxa presentes en el Pozo 5, sobre todo de las partes con mayor contenido cárnico. El Pozo 5 corresponde probablemente a un basurero de consumo de estas especies, ya que casi no se encontraron partes esqueléticas de poca carne (metapodios, falanges, vértebras caudales), que podrían haber sido resultado de un procesamiento de carnicería. Aunque existe la posibilidad que algunos elementos esqueléticos faltantes fueron seleccionados para la elaboración de artefactos, cabe mencionar que en ningún contexto del sitio se encontraron restos significantes de artefactos de hueso terminados o en proceso de elaboración.
Los materiales de la Estructura 5D2 consisten en abundantes restos de venados, así como diversas especies de reptiles medianos, todos estos restos de consumo. Los venados están representados prácticamente por todo el esqueleto, con huellas de preparación tanto en las extremidades como en los fragmentos del cráneo. A diferencia del Pozo 5 se encontraron aquí también metapodios y falanges proximales de venado, las últimas con huellas de masticación carnívora, lo que puede indicar que los venados llegaban enteros a la Estructura 5D2, para descuartizarlos y prepararlos allí mismo. Las falanges medias y distales faltan posiblemente, porque quedaban anexas a las pieles que se trabajaban (Pohl 1990:158). Fuera del venado no se encontraron otros animales «ricos en carne» como se habían visto en el Pozo 5. En vez de esto se evidencia un consumo de reptiles medianos. Tanto en los restos de iguana negra como de boa se encontraron huellas de preparación que desmientan una intrusión natural de estos restos y favorecen su interpretación como alimento, aunque se argumenta que las serpientes se utilizaron más bien en rituales (Carr 1995:2,6).
De igual modo se consumieron probablemente algunos bagres marinos evidenciados por restos craneales. Sin embargo, la presencia de peces en los contextos no se debe necesariamente interpretar como una utilización alimenticia fuerte de estas especies, porque tanto la espina dorsal de bagre marino como la vértebra perforada de manta raya comprueban más bien su utilización como utensilios y/o adornos. De este modo se encontraron pocos indicios de un comercio de peces marítimos de la costa cercana a Siho, tal como se reporta para sitios como Chichen Itza (Carr 1995:3), aún más distantes del mar.
Los habitantes de la Estructura 5D2 parecen haber enriquecido su alimentación cárnica con pequeñas especies, un rasgo reportado de muchos sitios del área Maya durante el fin del Clásico Tardío y Clásico Terminal. La diferencia entre los taxa del Pozo 5 y los de la Estructura 5D2 hacen pensar que la gente que formó el basurero del Pozo 5 tuvo posiblemente un mejor acceso a animales más carnosos y mejores que los habitantes de la Estructura 5D2, por lo que se podría especular sobre diferencias socio-económicas entre ambos contextos. En este sentido representan los habitantes de las Estructuras 5D13/14 posiblemente una élite de mayor rango que los habitantes de la Estructura 5D2, un dato que queda aún por refutar a través de la excavación de las estructuras adyacentes al Pozo 5.
Las capas del Clásico Terminal de la Estructura 5D2 revelaron únicamente pocos restos de venados, sin embargo todos quemados y con huellas de corte, indicando su consumo. En las capas del Clásico Terminal del Pozo 5 abundan los restos de perro, mientras que había poco venado cola blanca y muy poco jabalí. Muchos fragmentos de las capas correspondientes al Clásico Terminal no se pudieron identificar a más que como «mamífero mediano-grande» debido a su fuerte fragmentación. Estos fragmentos indeterminables proceden de diáfisis de huesos largos y son la única evidencia de huellas de preparación en forma de rupturas en fresco y marcas de fuego.
Tanto referente al Pozo 5 como a la Estructura 5D2, se postula un aprovechamiento intensivo de los pocos animales grandes, asando porciones de las extremidades (manchas negras en las superficies del hueso que al momento de asar no estuvieron cubiertas de carne), y luego ruptura de los huesos con el fin de obtener la médula (pequeños fragmentos de lascas óseas con fractura espiral como resultado de impactos para quebrar el hueso). Los fragmentos óseos de animales grandes del Clásico Terminal son más pequeños que en las capas del Clásico Tardío, lo que se interpreta aquí como una intensificación en su uso, junto con una reducción en la diversidad y cantidad de los taxa.
Los patrones que se evidencian en los materiales faunísticos de la Estructura 5D16 favorecen la interpretación mencionada anteriormente. También en la Estructura 5D16 se encontraron restos muy fragmentados de mamíferos medianos-grandes. También estos fragmentos proceden de diáfisis de huesos largos y muestran igualmente ruptura en fresco y huellas de fuego. Sin embargo, la cantidad de estos restos, en proporción a la totalidad de material faunístico recuperado en la estructura, es mucho menor y como complemento aparecen restos de roedores y reptiles medianos, muchos con huellas de preparación. Igualmente se encontraron restos de alas de aves, a lo contrario de lo que se esperaría normalmente, ya que las aves se representan más frecuentemente por las extremidades traseras (Emery 1997:313; Wing 1980:331), siendo éstas las partes más carnosas.
Los habitantes de la Estructura 5D16 tuvieron, al parecer, menos acceso a animales grandes que los habitantes del centro inmediato, que comprenden las Estructuras 5D2 y 5D13/14 (Pozo 5). Se podría suponer una diferencia en cuanto al poder en el sitio respecto al acceso a bienes de prestigio, de los cuales la carne, en tiempos de escasez, seguramente era uno. Las diferencias socio-económicas, y con esto las posibles diferencias en el poder, se expresan en la ubicación aislada de la Estructura 5D16 en cuanto al centro, así como respecto al hecho que la Estructura 5D16, a pesar de haber recibido una mayor inversión de trabajo en su construcción, está acompañada solamente por dos estructuras perecederas, mientras que las estructuras del centro yacen en la vecindad de la pirámide principal y de varios palacios.
CONCLUSIONES
Resumiendo, se puede decir que la fauna consumida del Clásico Tardío de Siho consiste principalmente en mamíferos grandes (venado cola blanca, jabalí y probablemente perro), así como en reptiles medianos (iguanas y tortugas; Figura 7), y representa grosso modo en el lado cárnico de la llamada «dieta tradicional Maya» del Clásico de las Tierras Bajas (entre otros Wing 1981:27). Especialmente la preponderancia del venado cola blanca es un rasgo muy común de esta época en toda Mesoamérica (Álvarez 1999; Carr 1995, 1996; Emery 1997; Pohl 1976, 1990; Wing 1980). Los habitantes de la Estructura 5D2 consumieron además reptiles y aves, aunque éstos no sobresalen en comparación con el venado.
Con el Clásico Terminal llega un cambio en los patrones alimenticios (Figura 8). Según las evidencias empiezan a reducirse los animales grandes en Siho a partir de este momento, un rasgo que se ve expresado en cuatro puntos.
- En primer lugar aparecen mucho menos restos de los taxa grandes en este periodo (venado cola blanca, jabalí).
- Estos restos estuvieron además quebrados a mayor escala, presentando casi todos fracturas en fresco, lo que indica que lo poco que se tenía fue aprovechado a mayor escala, rompiendo una multitud de huesos completamente con el fin de alcanzar el tuétano.
- En las capas terminales del Pozo 5 aumentan además mucho los restos del perro, animal domesticado cuyo consumo pudiera haberse visto como una ayuda inmediata contra la reducción de la fauna grande silvestre.
- La cuarta evidencia es la apariencia proporcionalmente alta de animales pequeños (roedores, reptiles), en la dieta de los habitantes de la Estructura 5D16 durante el Clásico Terminal, junto con escasos restos muy fragmentados en fresco de venado.
Podría argumentarse que la reducción de fauna grande en Siho durante el Clásico Terminal en comparación con el periodo anterior se debe a la reducción poblacional humana ocurrida entonces (Cobos et al. 2002). Sin embargo, una reducción poblacional frente a condiciones medio ambientales óptimas no hubiera requerido una intensificación de los medios de procuramiento.
En los análisis zoo-arqueológicos de Dzibilchaltun (Wing 1980), y Ceibal (Pohl 1990:167), se observó igualmente una tendencia de reducción de fauna grande, junto con un incremento de fauna pequeña en los contextos alimenticios del Clásico Tardío y Terminal. Los autores relacionan este fenómeno con una fuerte presión de cacería y deforestación durante el Clásico Tardío (Carr 1995:4; Wing 1980:331). Wing propone una reducción de fauna grande durante las fases tardías en Dzibilchaltun en comparación con el Preclásico. En Siho, la fauna grande parece haber estado presente aún durante el Clásico Tardío y la reducción fuerte inició al parecer durante el Clásico Terminal. Yula, en Yucatán, por ejemplo, a pesar de ser una muestra zoo-arqueológica relativamente pequeña, presenta perfiles faunísticos del Clásico Tardío-Terminal ricos en mamíferos grandes (Carr 1995:8), al igual que el área de Petexbatun, en donde no se registró una reducción de fauna grande hacia este periodo (Emery 1997:357).
Aún queda mucho por investigar en cuanto a los patrones de consumo cárnico del Clásico en las Tierras Bajas Mayas. La reducción de fauna grande por presión de cacería, deforestación y desplazamiento de las poblaciones silvestres, puede haber ocurrido durante distintas fases y en distintas áreas. A través de futuras investigaciones se espera poder investigar más a fondo cuáles fueron las causas de la postulada reducción de animales grandes en las Tierras Bajas Mayas del Norte, con qué repercusiones del proceso histórico Maya se relacionan, y por cuáles razones no se presentan en todos sitios al mismo tiempo. Estas investigaciones no solamente podrían aclarar muchos aspectos acerca de la vida de los Mayas prehispánicos, sino que pueden dar indicios hacia el importante tema de la conservación del medio ambiente actual.
AGRADECIMIENTOS
Se agradece a Rafael Cobos Palma, Lilia Fernández Souza y Socorro Jiménez Álvarez, por informaciones referentes al sitio. La investigación de campo en Siho durante 2001 y 2003 se efectuó gracias al apoyo otorgado por el Consejo de Arqueología del Instituto Nacional de Antropología e Historia, el Consejo Nacional de Ciencia y Tecnología (CONACYT-Proyecto 38092-H), el Programa de Naciones Unidas para el Desarrollo y la Universidad Autónoma de Yucatán.
REFERENCIAS
Álvarez, Ticul y Aurelio Ocaña
1999 Sinopsis de restos arqueo-zoológicos de vertebrados terrestres – Basada en informes del Laboratorio de Paleozoología del INAH. INAH Colección Científica 386. México.
Carr, Helen Sorayya
1995 Some Reflections on Interpreting Archaeofaunas from Northern Yucatan. Ponencia, 60 Reunión Annual, Society for American Archaeology, Minneapolis.
1996 Precolumbian Maya Explotation and Management of Deer Populations. En The Managed Mosaic: Ancient Maya Agriculture and Resource Use (editado por S. Fedick), pp.251-261. University of Utah Press, Salt Lake City.
Chase, Diane y Arlen Chase
2000 Inferences about Abandonment: Maya Household Archaeology and Caracol, Belize. Mayab 13:66-77.
Cobos, Rafael, Lilia Fernández Souza, Nancy Peniche May, Edgar D. Pat Cruz, Socorro Jiménez Álvarez, Vera Tiesler Blos, Christopher M. Götz y Alfonso Lacadena
2004 Proyecto Arqueológico El Surgimiento de la Civilización en el Occidente de Yucatán: Los Orígenes de la Complejidad Social en Siho. Informe de Actividades de la Temporada de Campo 2001 presentado al Consejo de Arqueología del INAH. Universidad Autónoma de Yucatán, Facultad de Ciencias Antropológicas, Mérida, Yucatán.
Cobos, Rafael, Lilia Fernández Souza, Vera Tiesler, Pilar Zabala, Armando Inurreta, Nancy Peniche May, Ma. Luisa Vázquez y Diana Pozuelo
2002 El Surgimiento de la Civilización en el Occidente de Yucatán: Los Orígenes de la Complejidad Social en Siho. Informe de Actividades de la Temporada de Campo 2001 presentado al Consejo de Arqueología del INAH. Universidad Autónoma de Yucatán, Facultad de Ciencias Antropológicas, Mérida, Yucatán.
Dunning, Nicholas P.
1993 Preliminary Data of the Site of Siho. Mecanuscrito. Archivo de la Sección de Arqueología del Centro INAH Yucatán, Mérida.
EA.FLMNH
2003 Colección fotográfica digital de 243 especímenes endémicos de la región Maya, efectuadas por Christopher Götz y en posesión del departamento de Arqueología Medio Ambiental del Florida Museum of Natural History, Vol. 2.
Emery, Katherine F.
1997 The Maya Collapse: A Zooarchaeological Investigation. Tesis doctoral, Anthropology, Graduate School, Cornell University, Ithaca.
Fernández Souza, Lilia, Marisa Vázquez de Agredos y Rafael Cobos Palma
2003 Análisis de una estructura de tipo palacio en Siho, Yucatán. En XVI Simposio de Investigaciones Arqueológicas en Guatemala, 2002 (editado por J.P. Laporte, H. Escobedo, B. Arroyo y H. Mejía), pp.1031-1036. Museo Nacional de Arqueología y Etnología, Guatemala.
Fernández Souza, Lilia y Nancy Peniche May
2004 Informe de la excavación horizontal realizada en la Estructura 5D2. En El Surgimiento de la Civilización en el Occidente de Yucatán: Los Orígenes de la Complejidad Social en Siho. Informe de Actividades de la Temporada de Campo 2001 presentado al Consejo de Arqueología del INAH (editado por R.Cobos Palma), pp.41-68. FCA UADY, Mérida.
Gautier, Achilles
1984 How Do I Count You? Let Me Count The Ways. Problems of Archaeozoological Cuantification. En Animals and Archaeology 4. Husbandry in Europe (editado por C. Grigson y J. Clutton-Brock), pp.237-251. British Archaeological Reports International Series 227, Oxford.
Gilbert, Miles B.
1980 Mammalian Osteology. Missouri Archaeological Society, Laramie, Wyoming.
Gilbert, Miles B., Larry D. Martin y Howard G. Savage
1985 Avian Osteology. Missouri Archaeological Society, Laramie, Wyoming.
Jiménez Álvarez, Socorro
2004 Siho como Unidad Política. Ponencia Congreso Internacional de Mayistas, Villahermosa, Tabasco.
Lacadena, Alfonso
2004 El Panel 1 de Siho: Informe epigráfico. En El Surgimiento de la Civilización en el Occidente de Yucatán: Los Orígenes de la Complejidad Social en Siho. Informe de Actividades de la Temporada de Campo 2001 presentado al Consejo de Arqueología del INAH (editado por R. Cobos Palma), pp.159-162. FCAUADY, Mérida.
Lee, Julian C.
2000 A Field Guide to the Amphibians and Reptiles of the Maya World. Cornell University Press, Ithaca.
Leopold, A. Starker
2000 Fauna Silvestre de México. Editorial Pax México. México D.F. (reimpresión de obra original de 1959).
Maler, Teobert
1997 Península Yucatán (editado por H. J. Prem). Monumenta Americana 5, Gebrüder Mann Verlag, Berlin.
O’Connor, Terry
2000 The Archaeology of Animal Bones. Texas A&M University Press, College Station.
Olsen, Stanley J.
1968 Fish, Amphibian and Reptile Remains from Archaeological Sites. Papers of the Peabody Museum of Archaeology and Ethnology, Vol. 56 (2), Harvard University, Cambridge, Massachusetts.
Peterson, Roger Tory y Edward L. Calif
1973 Mexican Birds. Peterson Field Guides, Houghton Mifflin Company, Boston & New York.
Pohl, Mary Ellen Deland
1976 Ethno zoology of the Maya: An Analysis of Fauna from Five Sites in the Peten, Guatemala. Tesis doctoral, Anthropology, Harvard University, Cambridge, Massachusetts.
1990 The Ethnozoology of the Maya. En Excavations at Seibal (editado por G. R. Willey), pp.142-174. Harvard University, Cambridge, Massachusetts.
Reid, Fiona A.
1997 A Field Guide to the Mammals of Central America and Southeastern Mexico. Oxford University Press, New York.
Tun Ayora, Gabriel Ernesto
2004 La organización de viviendas Mayas prehispánicas: Análisis de estructuras domésticas asociadas a unidades habitacionales de élite de Siho, Yucatán. Tesis de Licenciatura, Especialidad de Arqueología, Facultad de Ciencias Antropológicas, Universidad Autónoma de Yucatán, Mérida, México.
Wing, Elizabeth S.
1980 Appendix: Vertebrate Remains from Dzibilchaltun. En Excavations at Dzibilchaltun (editado por E. Andrews Wyllys IV y E. Wyllys Andrews V), pp.326-331. Middle American Research Institute, Tulane University, Publ.48. New Orleans.
1981 A Comparison of Olmec and Maya Foodways. En The Olmec and Their Neighbours (editado por E. Benson), pp.21-28. Dumbarton Oaks, Washington D.C.
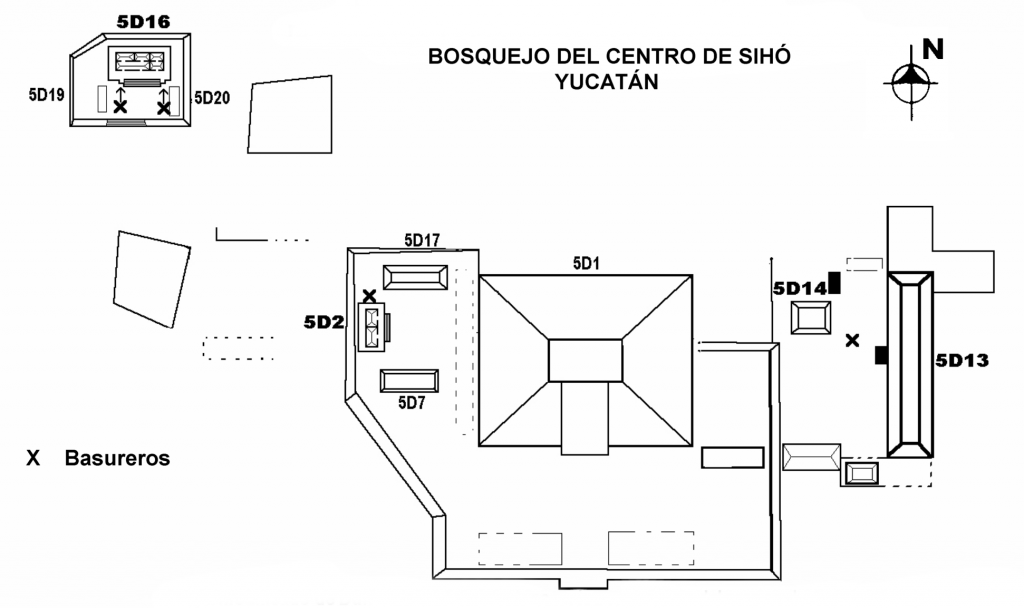
Figura 1 Bosquejo del centro de Siho, Yucatán, mostrando las estructuras mencionadas en el texto
Taxon
Cabeza
Esqueleto axial
Extremidades
Patas
NISP total
MNI por especie o género
n.d.
delanteras
traseras
superiores
inferiores
superiores
inferiores
5D2
Clásico Tardío
Bagre marino
1
1
1
Mantaraya
1
1
1
Reptil, n.d.
1
8
9
—
Iguana negra
9
2
3
3
17
4
Serpiente Boa
4
4
1
Serpiente Cascabel
1
1
1
Tortuga terrestre
2
2
1
Ave med.
2
2
—
Mamífero med.
7
7
—
Venado n.d.
12
1
8
1
1
23
—
Venado temazate
2
2
1
Venado cola blanca
11
1
3
3
6
24
2
Clásico
Terminal
Venado n.d.
1
1
—
Venado cola blanca
4
1
1
6
1
5D16
Clásico Terminal
Bagre marino
1
1
1
Iguana negra
1
1
2
1
Reptil peq. – med.
3
3
—
Serpientes
7
7
—
Ave med.
1
1
—
Roedor n.d.
14
14
—
Tuza
1
2
2
5
1
Perro
1
1
1
Mamífero med. – grd.
4
4
—
Venados
1
1
1
5D13 / 5D14 (Pozo 5)
Clásico Tardío
Pavo del monte
1
2
3
1
Perro
2
8
1
10
1
Mamífero med.
50
10
16
76
—
Venado n.d.
6
20
26
—
Venado cola blanca
3
5
16
6
30
4
Jabalí de collar
7
2
9
1
Clásico Terminal
Perro
17
2
19
1
Mamífero med.
3
15
18
—
Venado cola blanca
4
4
1
Jabalí de collar
1
1
1