

Descargar este artículo en formato PDF
Wright, Lori E.
1995 La dieta antigua y al evolución social de los Mayas del río Pasión: una visión isotópica. En VIII Simposio de Investigaciones Arqueológicas en Guatemala, 1994 (editado por J.P. Laporte y H. Escobedo), pp.258-268. Museo Nacional de Arqueología y Etnología, Guatemala (versión digital).
19
LA DIETA ANTIGUA Y LA EVOLUCIÓN SOCIAL DE LOS MAYAS DEL RÍO PASIÓN: UNA VISIÓN ISOTÓPICA
Lori E. Wright
Durante las últimas décadas, la arqueología ha empezado a desenmarañar la interacción entre las culturas prehistóricas y sus medio ambientes. Este progreso es especialmente visible en el caso de los Mayas de las Tierras Bajas, cuyo bosque tropical ha sido objeto de escrutinio intensivo. Como ya sabemos, las grandes poblaciones Clásicas fueron sostenidas por diversas formas de agricultura intensiva y, además, por medio de la explotación de una variedad de alimentos silvestres y domesticados. Las adaptaciones sociales también han sido aclaradas por estudios extensivos de los patrones de asentamiento y por el desciframiento de gran parte del sistema de escritura jeroglífica.
No obstante, la explicación dominante para el colapso de la civilización Maya en las Tierras Bajas es todavía ecológica y en su fundamento, Malthusiana. Se argumenta que las poblaciones aumentaron fuera de la capacidad del frágil ecosistema tropical (Culbert 1988; Santley et al. 1986). Dicho modelo propone que el aumento de la población causó una expansión de los sistemas agrícolas hasta el punto de utilizar todas las tierras disponibles. La erosión de los suelos, invasión de hierbas y la deforestación restringieron severamente la productividad agrícola en el Clásico Tardío. Frente a estos problemas, se argumenta que los agricultores se concentraron en la siembra de cosechas de uso corriente, especialmente de maíz. Estos cambios agrícolas transformaron la dieta consumida por los miembros de esta sociedad y ello implicó el deterioro del estado nutricional y de la salud de la población. A la larga, este cambio dietético se eslabona a la inestabilidad demográfica que ocasionó el abandono del área.
En su fundamento, el modelo es biológico y puede ser evaluado por información biológica de los restos óseos humanos. Este artículo examinará tres expectativas para la dieta prehistórica Maya que tienen su origen en este modelo ecológico. Para evaluarlas se utilizará información química obtenida de los huesos humanos recuperados en varios sitios de la región del río Pasión en el suroeste de Petén.
Aunque el modelo ecológico ha dominado las discusiones del Colapso Maya en el sentido general y teórico, el Colapso en la región del Pasión ha sido caracterizado frecuentemente por factores políticos. Trabajos en Altar de Sacrificios y Ceibal vislumbraron la posibilidad de que una invasión de extranjeros desde Campeche podría haber ocasionado el colapso de estos dos sitios (Sabloff y Willey 1967; Thompson 1970). Estudios epigráficos más recientes han demostrado que el Pasión llegó a ser un foco de intensa competencia política y militar entre unidades políticas locales al fin del periodo Clásico (Houston 1993). Las excavaciones del Proyecto Petexbatun ilustran que esta guerra causó el abandono de varios sitios cercanos a la Laguna Petexbatun (Demarest et al. 1989-1994). Esta nueva información política atisba la posibilidad de que factores sociales en vez de ecológicos podrían haber estado involucrados en la transición del Clásico Terminal.
EL ANÁLISIS ISOTÓPICO ÓSEO Y LA DIETA
El análisis químico de huesos proporciona información sobre la dieta prehistórica ya que la composición de los alimentos influye en la composición de los huesos. Básicamente: eres lo que comes. Hoy, enfocaré el análisis dietético por medio de los isótopos estables de carbono y nitrógeno. El elemento carbono, al igual que el nitrógeno, existe en varias formas de átomos, llamados isótopos, que se distinguen por el número de neutrones en el núcleo. Lo más abundante es el carbono-12 que contiene seis protones y seis neutrones. El carbono-13 contiene un neutrón extra. Pero en contraste al carbono-14 (con dos neutrones extra) que utilizamos para calcular la edad de rasgos arqueológicos, el carbono-13 es estable. Es decir que el núcleo no decae con el transcurso del tiempo. La cantidad de carbono-13 en un objeto orgánico depende de la cantidad ingerida por el organismo cuando estaba vivo. Igualmente, el nitrógeno-14 es el isótopo estable más abundante de este elemento, pero existe también el isótopo estable nitrógeno-15, que contiene un neutrón extra.
Para cuantificar la composición isotópica de una muestra nos referiremos a la relación entre las cantidades del isótopo más pesado y el menos pesado, es decir la relación entre carbono-13 y carbono-12 (ie.13C/12C), que designamos como «delta carbono-13», o sea «d13C». Los valores más positivos en la escala de d13C indican relativamente mayor contenido de 13C, el isótopo pesado. Decimos entonces que están enriquecidas con 13C o sea más pesadas. De manera similar, medimos la cantidad de nitrógeno-15 relativa al nitrógeno-14 (ie.15N/14N), designado como «d15N».
La proporción de los isótopos estables en los tejidos de un animal o planta depende de los procesos metabólicos y ecológicos. Por ejemplo, las plantas utilizan varias reacciones químicas en fotosíntesis. Plantas que utilizan la vía C4 incorporan más carbono-13 que los que utilizan la vía C3, porque usan una enzima distinta con mayor afinidad al carbono-13. Aquí en Guatemala, el maíz es una de las pocas plantas comestibles del tipo C4 y muestra una composición isotópica cercana a -10 por mil. La mayoría de las otras plantas utilizan la vía fotosintética C3, con una composición de -26%o.
Un herbívoro tendrá tejidos con una composición isotópica parecida a las plantas de su dieta. Entonces, los animales que consumen mucho maíz se caracterizan por un d13C pesado, semejante al maíz. Los que comen otras plantas se caracterizan por d13C bajo. Además, hay un factor de enriquecimiento isotópico entre la comida y los tejidos. Para el carbono, estimamos que el colágeno de los huesos está enriquecido un 5%o sobre la composición media de la dieta.
En el caso de nitrógeno, las plantas que contienen bacterias simbióticas en las raíces, tal como el frijol, muestran niveles de d15N un poco más bajo que la mayoría de las plantas que obtienen su nitrógeno del suelo. Todos los animales obtienen nitrógeno solamente de proteínas dietéticas. Su colágeno está enriquecido al 3%o sobre la fuente dietética.
Así, el análisis isotópico del colágeno óseo presenta un instrumento poderoso para examinar el consumo del maíz y la carne, dos alimentos significativos en el modelo ecológico del colapso Maya.
LAS EXPECTATIVAS DEL MODELO ECOLÓGICO
En varios trabajos (Santley, Killion y Lycett 1986; Willey y Shimkin 1973; Sanders 1962, 1963; Culbert 1988), entre otros, el modelo atribuye el deterioro posterior del medio ambiente y de la salud al cultivo del maíz. Por lo tanto, el análisis isotópico debería confirmar que los pueblos del Pasión consumieron el maíz como su principal fuente nutritiva. Segundo, en esta propuesta debe haber aumentado el consumo de maíz durante la ocupación tardía, con los agricultores produciendo un mayor cultivo de este grano durante los periodos de presión poblacional. Esta desviación hacia el maíz debería estar reflejada en un incremento en la tasa isotópica de carbono en los huesos humanos. Tercero, el medio ambiente selvático que fue una fuente de caza, disminuyó durante el Clásico Tardío debido a la expansión de los campos agrícolas hacia todos los terrenos cultivables. A la par, el incremento en la caza por el aumento de la población humana limitó la población de animales salvajes, hasta que las provisiones de carne fueron muy restringidas. Químicamente, esto se identificará por un declive en la tasa isotópica de nitrógeno en relación al tiempo en el colágeno humano.
MÉTODOS ANALÍTICOS Y LA PRESERVACIÓN ÓSEA
Se tomaron muestras de hueso cortical de todos los esqueletos adultos bien preservados de los sitios de Altar de Sacrificios, Ceibal y Dos Pilas. También se analizaron algunos huesos arqueológicos de animales recuperados en las excavaciones del Proyecto Petexbatun. Los análisis isotópicos se llevaron a cabo en los laboratorios de la Dra. Schoeninger en la Universidad de Wisconsin y del Dr. Tieszen en Augustana College. Se limpiaron los pedazos de hueso con agua destilada y se descalcificaron en EDTA. La composición isotópica de carbono y nitrógeno del colágeno fue medida por espectrometría de masas.
A pesar de la mala conservación ósea en este ambiente húmedo, en general la preservación del colágeno era buena. De las 121 muestras humanas preparadas solamente nueve no produjeron un residuo orgánico y no fue posible analizarlas. La relación de C/N y el contenido de C y N fueron medidos para investigar el estado del colágeno en las muestras restantes. La mayoría de hueso del Pasión muestra C/N entre el rango esperado para el colágeno, entre 2.9 y 3.6 (DeNiro 1985). Todos contenían aproximadamente 45% de carbono y 16% de nitrógeno, que es semejante al colágeno de tendones bovinos (Ambrose 1990). Ocho muestras se eliminaron del análisis estadístico porque tenían C/N elevados o valores isotópicas fuera del rango de los demás, de una muestra humana de 104 esqueletos en total.
RESULTADOS ISOTÓPICAS Y LA DIETA PREHISTÓRICA
La distinción isotópica entre el maíz y las plantas C3 provee la base para identificar el consumo de maíz (Figura.1). Pero dos plantas, el cacto nopal (Opuntia spp.) y la piñuela (Bromelia karatas) usan la vía fotosintética CAM (Crassulacean Acid Metabolism) y muestran d13C similar al maíz. Su consumo puede contribuir al enriquecimiento del colágeno humano.
La mayoría de los venados, jabalíes y tepezcuintes ilustran delta carbono bajo indicando que consumieron principalmente plantas C3 selváticas. Pero, dos venados arqueológicos de Aguateca y dos pecaríes modernos fueron más pesados en d13C, lo que indica que invadían las milpas para comer maíz. Como el área de Petexbatun es un área fronteriza hoy en día y la densidad de la población moderna es menor que lo hipotetizado para el periodo Clásico, este resultado enriquecido es notable. El d13C del pecarí implica que la mitad del carbono ingerido por este animal se originó de plantas C4. A la inversa, el d13C bajo de la mayoría de herbívoros arqueológicos implica que un área substancial de bosque fue mantenido en la región durante el Clásico Tardío.
Un resultado interesante es la composición baja en 13C de los peces de la Laguna Petexbatun. Además, los peces muestran d15N pesado por su dieta carnívora. Entre los carnívoros, analicé muestras de un puma o jaguar arqueológico, un tigrillo moderno, un cocodrilo arqueológico y varias culebras modernas. Para carbono y nitrógeno, los carnívoros presentan enriquecimiento trófico sobre los valores de los herbívoros, siendo más positivo o pesado. Es importante notar que los gatos ilustran d15N en el rango de las muestras humanas. Además, el felino arqueológico se acerca a los valores humanos en carbono, sugiriendo que ha consumido presas que a su vez comieron plantas C4.
El colágeno humano arqueológico es un poco más positivo en d13C que el maíz, como es de esperar si el maíz era una comida importante y de uso corriente. Asimismo, los resultados pesados en d13C en los dos venados arqueológicos reconfirman el papel importante del maíz en la agricultura prehistórica del Pasión. Sin embargo, la composición isotópica de nitrógeno de las muestras humanas es un poco pesada, semejante a la de los especímenes carnívoros. En verdad, el colágeno humano está en donde se espera encontrar el de un animal que ha consumido solamente carne de animales herbívoros. Por lo tanto, la primera expectativa del modelo ecológico del colapso solamente está apoyada parcialmente. El maíz era una comida importante para los Mayas del río Pasión. Pero el consumo de carne también era importante y contribuyó con una cantidad substancial de proteína dietética para la síntesis del colágeno.
Cambios cronológicos en el consumo del maíz y de la carne pueden ser evaluados con la información isotópica de cada sitio. Primero, en Altar de Sacrificios hay una tendencia hacia d13C más positiva desde el periodo Preclásico hasta el Clásico Tardío (Figura.2). En el Clásico Terminal, los habitantes de Altar de Sacrificios dejaron de construir edificios ceremoniales y dedicar monumentos jeroglíficos, pero una gran población quedó en el sitio. El d13C bajó un poco hacia el nivel del Clásico Temprano. Dado que los isótopos de nitrógeno no cambiaron durante estas épocas, es posible identificar estos cambios en d13C como desviaciones en la importancia relativa del maíz.
En Dos Pilas, una ocupación pequeña del Clásico Terminal data de una época posterior al retiro de la dinastía de Petexbatun hacia Aguateca y el abandono de la mayor parte del sitio. Cuatro esqueletos de esta comunidad remanente ilustran d13C un poco inferior a los entierros del Clásico Tardío, pero d15N equivalente a la época del apogeo del sitio (Figura.3). Esto sugiere una desviación hacia menor consumo de maíz después del colapso de la autoridad política en Dos Pilas, pero ningún cambio en los sistemas de obtención de proteína.
En contraste, el colágeno de Ceibal no muestra cambios en d13C durante la larga ocupación del sitio (Figura.4). La muestra pequeña de esqueletos Preclásicos tiene tasas isotópicas de carbono equivalentes al de los residentes tardíos. Estos son substantivamente más pesados que los esqueletos contemporáneos de Altar de Sacrificios, lo que puede indicar un mayor consumo de maíz en Ceibal. Además, tanto el colágeno Preclásico como el Clásico Tardío de Ceibal es isotópicamente más pesado en nitrógeno que el de Altar de Sacrificios. Esta diferencia química es debida al mayor consumo de carne en Ceibal. Puesto que los dos sitios se localizan entre zonas ecológicas semejantes, la discrepancia debe ser el resultado de preferencias culturales en lugar de determinantes agrícolas. Las muestras se diferencian en que los entierros de Ceibal fueron excavados en grupos domésticos y los de Altar de Sacrificios incluyen contextos de arquitectura pública. Pero la distinción no está de acuerdo con un modelo simple del consumo de carne y status social.
Los huesos del Clásico Tardío y Terminal de Ceibal muestran
niveles equivalentes de carbono, indicando estabilidad en la importancia del maíz en la ocupación del Clásico Terminal. En lugar de ello, el colágeno del Clásico Terminal muestra un declive en d15N relativo al Clásico Tardío. Esto puede indicar un declive en el consumo de carne en la ocupación final, de acuerdo con la expectativa del modelo ecológico del colapso. Pero, el colágeno del Clásico Tardío en Ceibal es el más alto en d15N de la región, siendo uno por mil más pesado que las dietas de Altar de Sacrificios o Itzan de cualquier época. La media del Clásico Terminal es bastante pesada, implicando un amplio consumo de carne. Hipotetizo que este cambio refleja un trueque de peces por plantas C3 —quizá frijol— en el Clásico Terminal, sin ningún cambio en el consumo de carne o de maíz. Sin embargo, el pequeño descenso en d15N del Clásico Terminal en Ceibal no implica una escasez crítica de proteína animal.
El contexto del entierro presenta una buena oportunidad para examinar aspectos sociales, porque el funeral es un rito muy importante en la vida de la persona y la posición social está expresada por los ritos practicados. Para examinar la variabilidad mortuoria, se utilizó el análisis de los componentes principales con variables binarias de la ubicación del entierro, la forma de sepultura, la orientación, la posición del esqueleto y las ofrendas. Estas manipulaciones estadísticas manifiestan una dimensión supraordinal de símbolos funerarios redundantes pan-Mayenses. Estos símbolos probablemente identifican los entierros de los personajes de la realeza. Para definir grupos sociales entre los entierros de cada sitio y periodo, se utilizó un análisis de agrupación de eslabonamiento completo (Complete Linkage Cluster Analysis). Diferencias dietéticas se encuentran entre estos grupos en el Clásico Tardío.
En Altar de Sacrificios, los dos esqueletos del grupo supraordinal o real se presentan con alto d15N, indicando un consumo diferencial de carne o peces (Figura.5). Se encuentra un patrón diferente en Dos Pilas, donde los dos entierros supraordinales tienen composiciones isotópicas distintas (Figura.6). El colágeno del Gobernante 2 de Dos Pilas es semejante al de los entierros sencillos del Grupo C con pocas ofrendas. El de su nuera, la Señora de Cancuen, es más similar al de los entierros de posición social intermedia, depositadas en criptas de caliza. Estos esqueletos en criptas con ofrendas y posiciones diferentes muestran d15N equivalentes, pero se distinguen por d13C. En cada sitio del Clásico Tardío, tanto los esqueletos masculinos como los femeninos fueron enterrados en cada tipo de sepultura y hay pocas diferencias sistemáticas entre los sexos. La posición social era la determinante más importante de la dieta del Clásico Tardío y no el sexo de la persona.
En contraste, patrones mortuorios relacionados con la dieta están ausentes en las grandes muestras de entierros del Clásico Terminal en Altar de Sacrificios y en Ceibal. La ausencia de diferencias dietéticas entre grupos mortuorios corre a la par de una falta de organización formal en el patrón mortuorio de los dos sitios. En lugar de un patrón social, las dietas del periodo Clásico Terminal muestran una división de sexos, con el colágeno de esqueletos masculinos más pesado en d13C que los femeninos (Figura.7). Los hombres del Clásico Terminal consumieron más maíz que las mujeres. Juntos, estos hallazgos vislumbran un cambio fundamental en la definición de posición socio-económica durante el Clásico Terminal.
APRECIACIONES GENERALES
Los resultados isotópicos no apoyan las tres expectativas del modelo ecológico. El maíz era una cosecha importante en la región del río Pasión, pero su consumo no se incrementó de manera uniforme con la expansión agrícola. Si el consumo de maíz era culpable de los problemas de salud que se discuten frecuentemente con respecto al colapso, los declives en d13C durante el Clásico Terminal implican un mejoramiento de salud y contradicen el modelo ecológico. Los resultados isotópicos de nitrógeno confirman que las provisiones de carne animal no fueron diezmadas por el incremento de la población humana. El análisis de los huesos de animales de Ceibal (Pohl 1985) y de los sitios de Petexbatun (Emery 1991), indican la explotación continua de la fauna silvestre y de manera indirecta, el mantenimiento de los bosques. Asimismo, los trabajos paleoecológicos de Dunning (comunicación personal, 1994) en la Laguna Tamarindito documentan poca erosión de suelo durante el Clásico Tardío pero sí la presencia de cantidades de polen de árboles selváticos. Aunque son preliminares, estos resultados indican que la deforestación no llegó a un punto crítico en el área de Petexbatun. Por otra parte, mis estudios del desarrollo dental, anemia e infección ósea no identifican ningún deterioro de la salud en el Pasión a través del tiempo (Wright 1994a, 1994b). En resumen, no hay evidencia concreta que apoye un modelo ecológico para el colapso de esta región.
Es importante recordar que el modelo ecológico se originó en una época cuando el mito de la milpa de maíz dominó nuestras percepciones de la agricultura prehistórica Maya. Ahora sabemos que los Mayas aprovecharon sistemas agrícolas complejos y que explotaron la diversidad microambiental (Atran 1993; Nations y Nigh 1980). Tenemos que ser cautelosos al elaborar analogías directas que proyectan las dietas Mayas actuales hacia atrás en la prehistoria. Además de quinientos años de cambio cultural Postclásico, la conquista española inició una transformación enorme en las prácticas dietéticas del Nuevo Mundo. Por ejemplo, fue hasta hace poco que la tortilla llegó ser un elemento omnipresente en las comidas Mayas. Como está representado en su arte, los Mayas de periodo Clásico consumieron maíz en forma de tamales, no de tortillas (Taube 1989). Hoy en día, el acceso al menú se relaciona más con la desigualdad política que con la preferencia dietética.
La información química y mortuoria ilumina posibles cambios en la organización social de la civilización Maya del Clásico Terminal. Los estudios epigráficos y arqueológicos están documentando una intensa competencia política entre las entidades políticas locales en la región del Pasión hacia el fin del periodo Clásico, lo que implica un cambio dramático en la interacción social. En lugar de la catástrofe ecológica, estas transformaciones sociales son la clave para desenmarañar el proceso del colapso.
AGRADECIMIENTOS
Este estudio se emprendió con el generoso permiso del Instituto de Antropología e Historia de Guatemala y del Museo Peabody de la Universidad de Harvard. El análisis químico fue posible por el apoyo financiero de las becas predoctorales extendidas a J. Buikstra y L. Wright por la Fundación Nacional de Ciencias de los Estados Unidos y de la Fundación Wenner-Gren de Investigaciones Antropológicas. Durante la ejecución del estudio recibí ayuda financiera del Consejo Nacional de Ciencias Naturales e Ingeniería de Canadá, la Federación Canadiense de Mujeres Universitarias y la Universidad de Chicago. También se agradece al Proyecto Arqueológico Regional Petexbatun y la Universidad de Vanderbilt su apoyo y su colaboración.
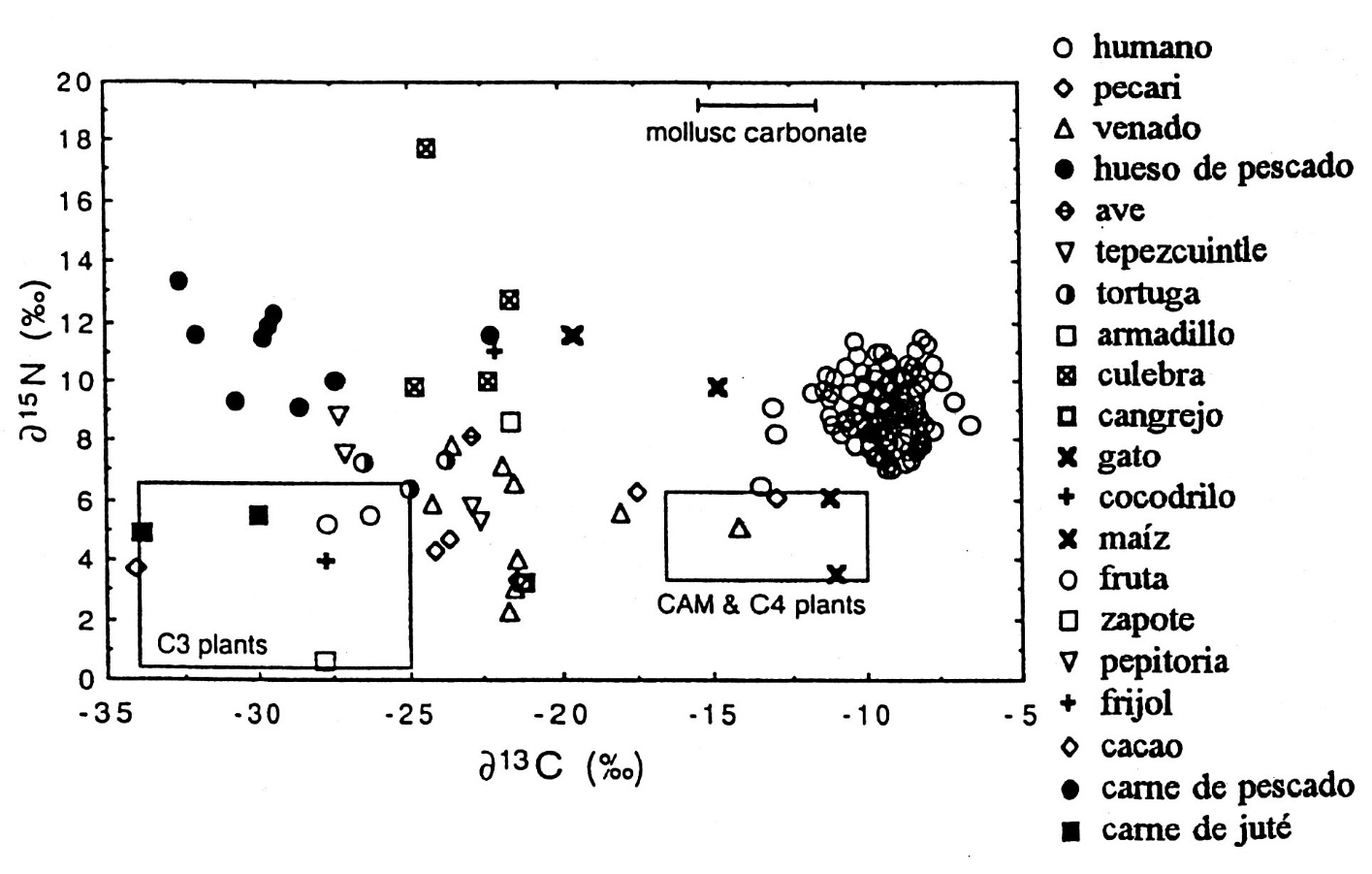
Figura 1
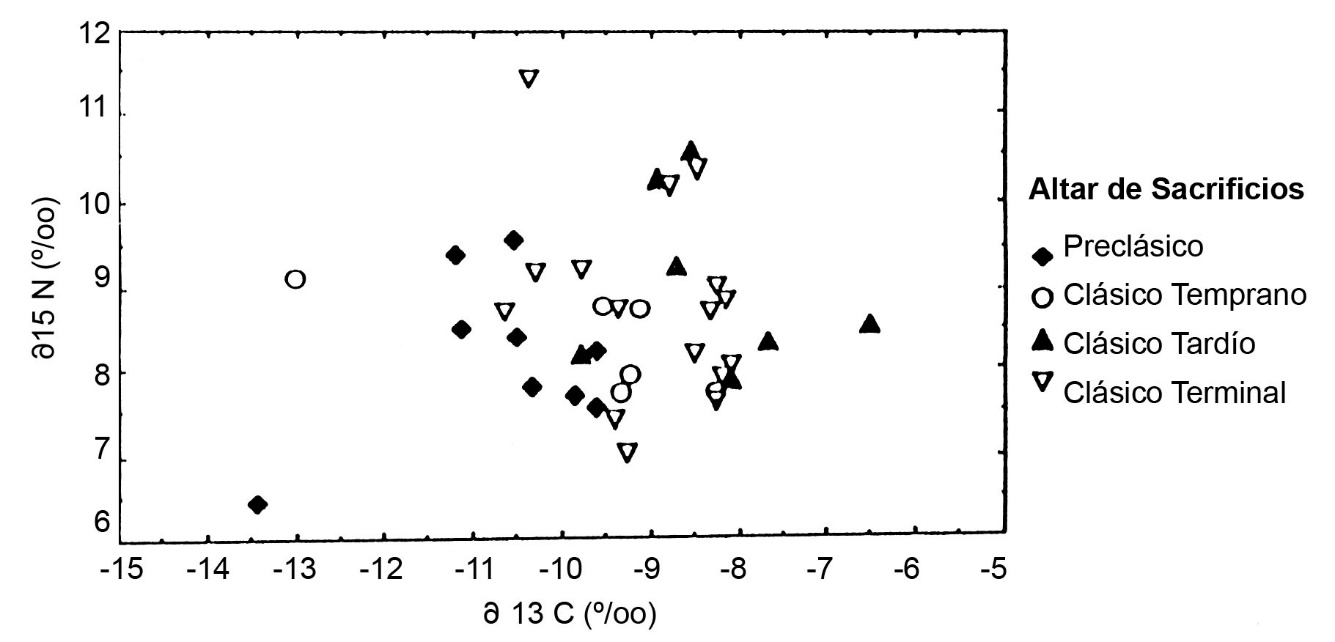
Figura 2
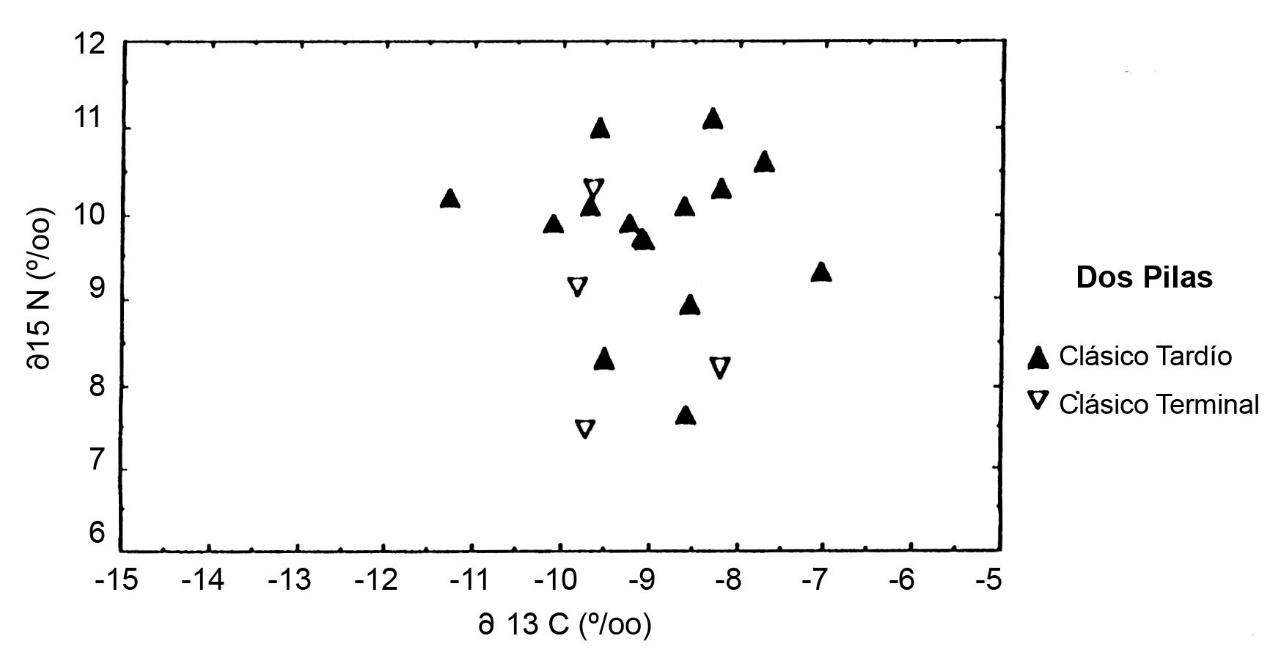
Figura 3
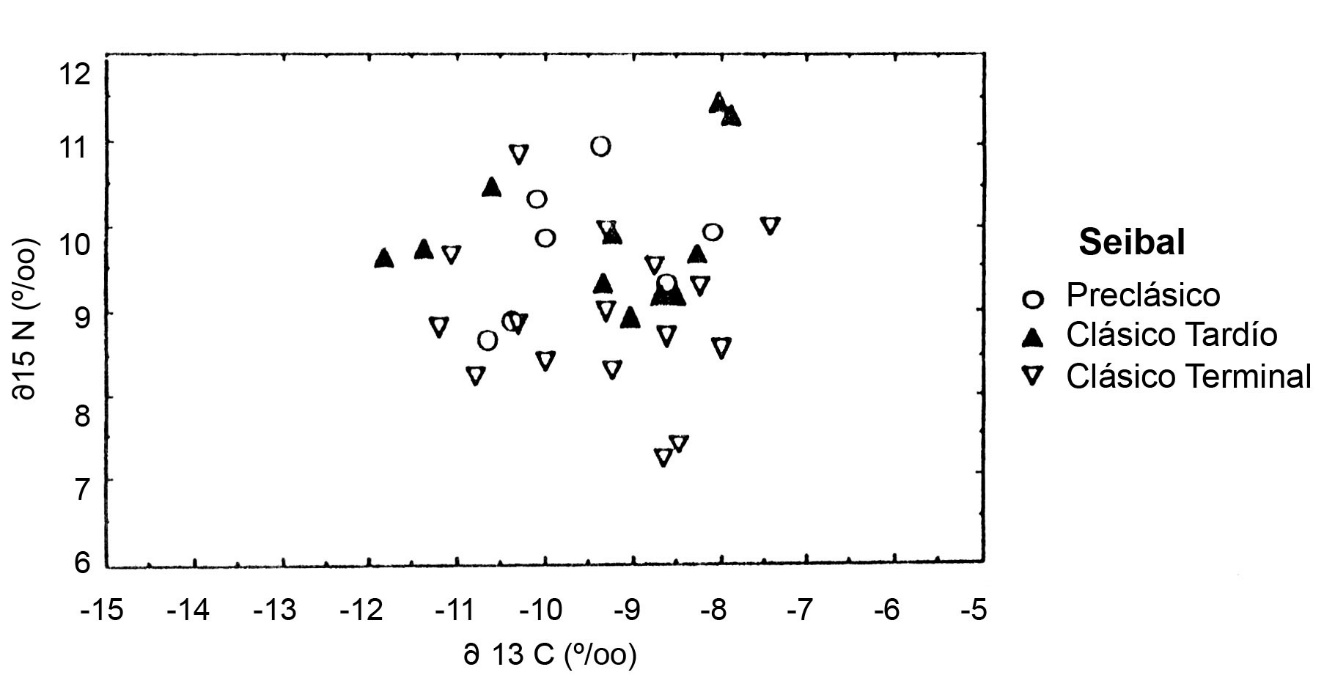
Figura 4
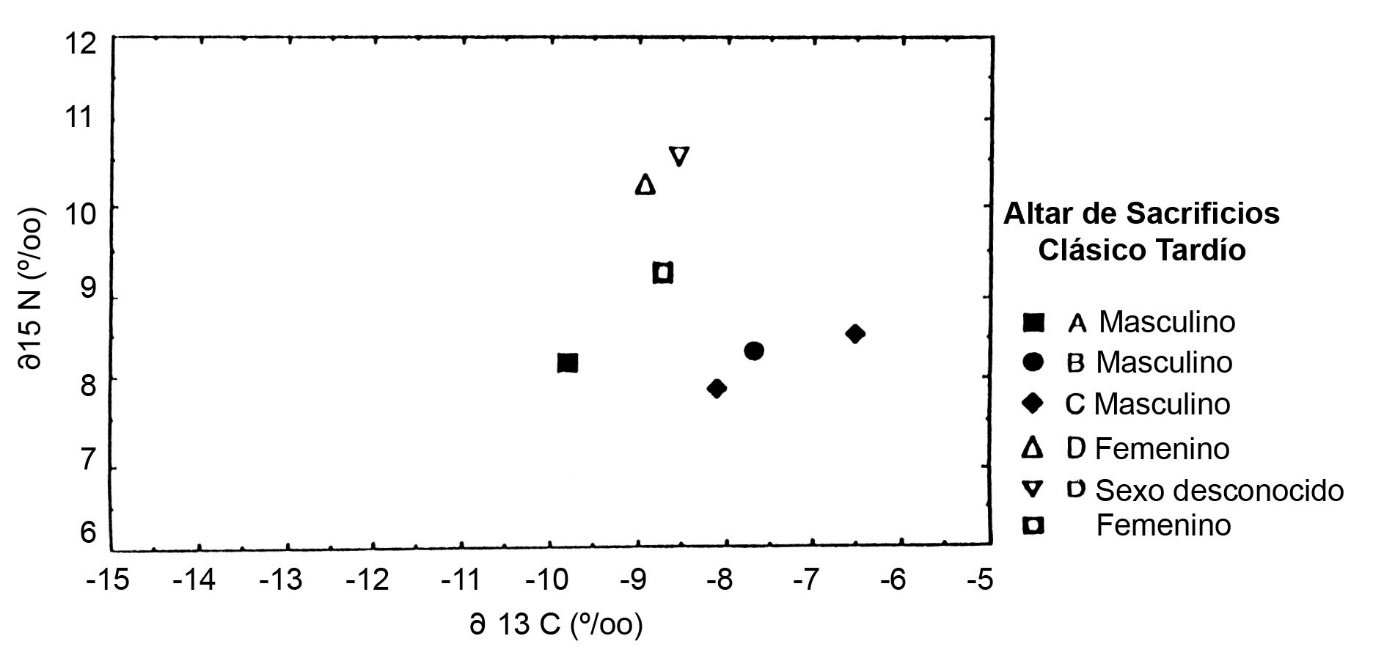
Figura 5
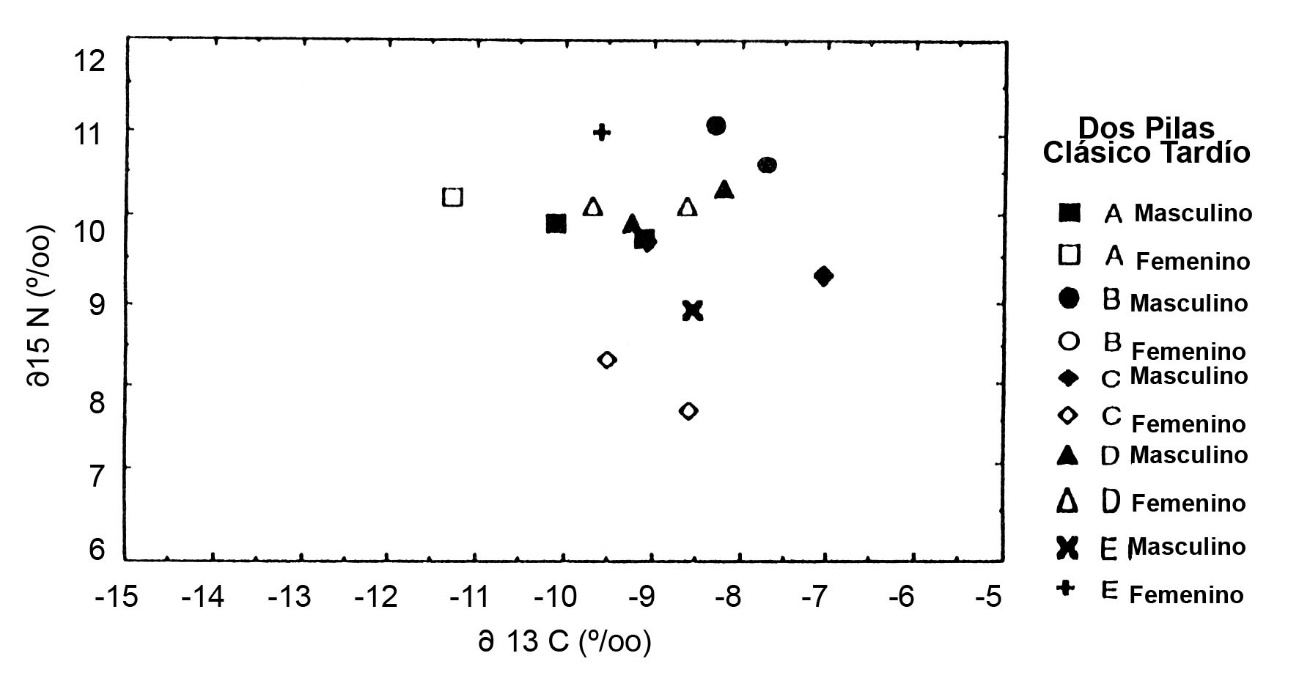
Figura 6
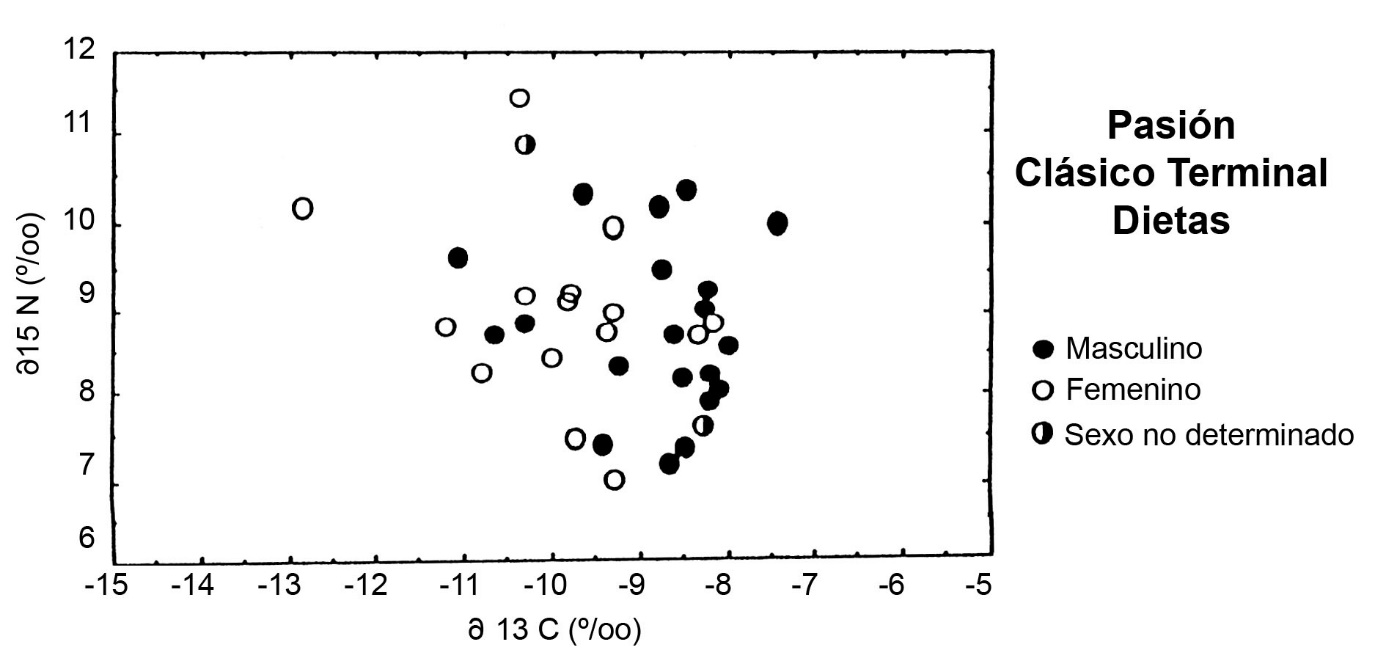
Figura 7
REFERENCIAS
Ambrose, S.H.
1990 Preparation and Characterization of Bone and Tooth Collagen for Isotopic Analysis. Journal of Archaeological Science 17:431-451.
Atran, S.
1993 Itza Maya Tropical Agro-Forestry. Current Anthropology 34:633-700.
Culbert, T. Patrick
1988 The Collapse of Classic Maya Civilization En The Collapse of Ancient States and Civilizations (editado por N. Yoffee y G.L Cowgill):69-101. University of Arizona Press, Tucson.
Demarest, Arthur A.
1989 The Olmec and the Rise of Civilization in Eastern Mesoamerica. En Regional Perspectives on the Olmec (editado por R.J. Sharer y D. Grove):303344. School of American Research Advanced Seminar Series, Cambridge University Press, Cambridge.
Demarest, Arthur A., H. Escobedo, J.A. Valdés, S. Houston, L. Wright y K. Emery
1991 Arqueología, Epigrafía y el descubrimiento de una tumba real en el centro ceremonial de Dos Pilas, Petén, Guatemala. U Tz’ib 1 (1):14-28. Asociación Tikal, Guatemala.
Demarest, Arthur A. et al. (ed)
1989-93 Proyecto Arqueológico Regional Petexbatun, Informes Preliminares Nos. 1-5. Proyecto Arqueológico Regional Petexbatun, Vanderbilt University, Nashville.
Demarest, Arthur A. y Takeshi Inomata
1992 Conclusiones Generales para la Temporada 1992 del Proyecto Arqueológico Regional Petexbatun. En Proyecto Arqueológico Regional Petexbatun: Informe Preliminar No.4 (A.A. Demarest, T. Inomata y H. Escobedo):370-383. Proyecto Arqueológico Regional Petexbatun, Vanderbilt University, Nashville.
Demarest, Arthur A. y Juan Antonio Valdés
1994 Proyecto Arqueológico Regional Petexbatun: Temporada 1993. En VII Simposio de Investigaciones Arqueológicas en Guatemala, 1993 (editado por J.P. Laporte y H.L. Escobedo):507-512. Museo Nacional de Arqueología y Etnología, Guatemala.
DeNiro, M.J.
1985 Postmortem Preservation and Alteration of in Vivo Bone Collagen Isotope Ratios in Relation to Palaeodietary Reconstruction. Nature 317:806-809.
Emery, Kitty
1991 Análisis Preliminar de los Restos Óseos de Fauna de la Región de Petexbatun. En Proyecto Arqueológico Regional Petexbatun, Informe Preliminar No.3, Vol.2 (A.A. Demarest, T. Inomata, H. Escobedo y J. Palka):813-828. Proyecto Arqueológico Regional Petexbatun, Vanderbilt University, Nashville.
Nations, James D. y R.B. Nigh
1980 The Evolutionary Potential of Lacandon Maya Sustained-Yield Tropical Forest Agriculture. Journal of Anthropological Research 36:1-30.
Pohl, M.
1985 The Privileges of Maya elites: Prehistoric Vertebrate Fauna from Seibal. En Prehistoric Lowland Maya Environment and Subsistence Economy (editado por M. Pohl):133-145. Papers of the Peabody Museum of Archaeology and Ethnology, Vol.77. Harvard University Press, Cambridge.
Sabloff, Jeremy A. y Gordon R. Willey
1967 The Collapse of Maya Civilization in the Southern Lowlands: A Consideration of History and Process. Southwestern Journal of Anthropology 23:311-336.
Sanders, William T.
1962-3 Cultural Ecology of the Maya Lowlands. Estudios de Cultura Maya 2: 79-121; 3:203-241.
Santley, S.R., T.W. Killion y M.T. Lycett
1986 On the Maya Collapse. Journal of Anthropological Research 42:123-159.
Taube, Karl Andreas
1989 The Maize Tamale in Classic Maya Diet, Epigraphy and Art. American Antiquity 54 (1):31-51.
Thompson, J. Eric S.
1970 Maya History and Religion. University of Oklahoma Press, Norman.
Wright, Lori E.
1993 Estudios Osteológicos en la Región de Petexbatun: Reporte Interno IV. En Proyecto Arqueológico Regional Petexbatun: Informe Preliminar No.5 (editado por J.A. Valdés, A. Foias, T. Inomata, H. Escobedo y A.A. Demarest):151-160. Proyecto Arqueológico Regional Petexbatun, Vanderbilt University, Nashville.
1994a The Sacrifice of the Earth? Diet, Health, and Inequality in the Pasion Maya Lowlands. Tesis Doctoral, Deptartment of Anthropology, University of Chicago, Chicago.
1994b Enfermedad, Salud y el Colapso Maya en las Tierras Bajas. En VII Simposio de Investigaciones Arqueológicas en Guatemala, 1993 (editado por J.P. Laporte y H. Escobedo):553-562. Museo Nacional de Arqueología y Etnología, Guatemala.
Willey, Gordon R. y D.B. Shimkin
1973 The Maya Collapse: A Summary View. En The Classic Maya Collapse (editado por T.P. Culbert):457-502. University of New Mexico Press, Albuquerque.