

Descargar este artículo en formato PDF
Scherer, Andrew K.
2006 Las relaciones biológicas durante la época Clásica Maya. En XIX Simposio de Investigaciones Arqueológicas en Guatemala, 2005 (editado por J.P. Laporte, B. Arroyo y H. Mejía), pp.595-604. Museo Nacional de Arqueología y Etnología, Guatemala (versión digital).
54
LAS RELACIONES BIOLÓGICAS
DURANTE LA ÉPOCA CLÁSICA MAYA
Andrew K. Scherer
Palabras clave
Arqueología Maya, Tierras Bajas, Guatemala, afinidad biológica de poblaciones, análisis estadísticos, estudios dentales
BIOLOGICAL RELATIONSHIPS OF THE CLASSIC MAYA
Much of our knowledge of the interaction between the Classic Maya sites is based on patterns of material culture and events documented through epigraphy. This paper presents a new line of evidence based on the genetic information from Classic Maya skeletons. Teeth were collected from 741 skeletons from 12 sites in Guatemala, Mexico, Belize and Honduras. These sites include Tikal, Calakmul, Piedras Negras, Copan, Kaminaljuyu, and sites from the Pasion River region and the Pacific Coast, among others. This study tests the hypothesis that the Classic Maya sites, located close to one another, show stronger biological relationships. In reality this is not the case. In fact, during the Classic period there was a high level of gene-flow between sites near and far away. These results are not surprising given the archaeological and epigraphic evidence for long distance interaction between Maya sites during the Classic period.
En la arqueología, el estudio de la historia de la población, encierra un amplio rango de preguntas de investigación que incluyen demografía, orígenes de la población y deceso de la misma, así como la interacción entre diferentes poblaciones. Con el propósito de reconstruir la historia de la antigua población Maya, tradicionalmente los investigadores han confiado en una combinación de datos epigráficos y material cultural, así como en estilos cerámicos y arquitectónicos. Sin embargo, los datos biológicos tomados de los restos de los Mayas en sí, también pueden ser una herramienta poderosa para reconstruir la historia de la antigua población.
En este informe se evaluará el conocimiento de las dinámicas de población durante el Clásico Maya a la luz de datos genéticos tomados directamente de antiguas osamentas Mayas. Desafortunadamente, debido a la pobre preservación de las mismas, por el momento no es posible extraer ADN de una manera confiable y consistente de esas osamentas, a pesar de los múltiples esfuerzos que se han realizado (Merriwether et al. 1997). Sin embargo, es posible usar fenotipos óseos y dentales para inferir información genética subyacente. Para este estudio se usaron medidas de los dientes, los cuales tienen características altamente hereditarias (Kieser 1990; Townsend y Brown 1978), como una herramienta para reconstruir las dinámicas de población de los antiguos Mayas.
Esta investigación inició con una simple hipótesis: probar si una población del Clásico localizada geográficamente cerca de otra, también exhibía afinidades biológicas fuertes, mientras que aquellas que se localizaban más apartadas unas de las otras, estarían menos relacionadas. En genética de poblaciones, esa situación se conoce como aislamiento por distancia. Por lo tanto, si el aislamiento por distancia resulta correcto para el Clásico Maya, se podría esperar que los sitios arqueológicos localizados en la región del río Usumacinta estén más relacionados los unos con los otros, más de lo que podrían estarlo con los sitios del centro de Petén.
Puede parecer intuitivo que las poblaciones que se localizan cerca las unas con las otras demostraran afinidades fuertes, tanto con un antepasado en común, como de un flujo genético en curso. Sin embargo, existe abundante evidencia arqueológica y epigráfica que sugiere que podría no ser ese el caso para el Clásico Maya. Eso se hace evidente, en especial, cuando se considera la dinámica política durante el Clásico Tardío, la cual se enfocaba alrededor de los sitios de Tikal y Calakmul. Ambos, Tikal y Calakmul, se localizan cerca el uno del otro, en el corazón de las Tierras Bajas Mayas, habiendo sido los dos sitios más grandes y más densamente poblados durante el Clásico (Culbert et al. 1990; Folan et al. 1995). Sin embargo, estas dos entidades políticas estuvieron muy lejos de ser vecinos amigables, teniendo regularmente luchas la una con la otra, tanto directa como indirectamente a través de sitios más pequeños que estuvieron aliados con esas superpotencias (Martin y Grube 1995).
Una de las mejores ilustraciones de la naturaleza de ese esfuerzo, proviene del sitio de Dos Pilas. En sus inicios, Dos Pilas fue fundado por la dinastía de Tikal en el año 629 DC, pero en el año 650 DC Calakmul saqueó Dos Pilas y desde entonces, Dos Pilas luchó del lado de Calakmul en contra de Tikal (Fahsen 2003). Con eso en mente, se exploró no sólo si el aislamiento por distancia explica la estructura de la población de la época Clásica Maya, sino también se investigaron los efectos de la hegemonía de Tikal y Calakmul sobre la historia de la población durante el Clásico Maya.
METODOLOGÍA
Con el objeto de estudiar relaciones biológicas en el área Maya, se recolectaron datos métricos dentales de 741 esqueletos provenientes de 12 sitios Mayas (Figura 1). Se seleccionaron estos 12 sitios debido a que tienen grandes colecciones óseas, que juntas representan siete diferentes zonas arqueológicas dentro del área Maya. En este informe se usarán las zonas arqueológicas que en un inicio fueron definidas por Culbert (1973), y que aún se continúan usando (Rice y Forsyth 2004). Las zonas arqueológicas proveen un marco adecuado para probar la hipótesis de aislamiento por distancia. Si el aislamiento por distancia explica la estructura de la antigua población Maya, se podría esperar una mayor similitud biológica entre los sitios de una misma zona arqueológica, que entre sitios de diferentes zonas. Esta investigación se enfoca en sitios del sur de las Tierras Bajas Mayas. Sin embargo, también se incluyó el sitio de Kaminaljuyu y algunos sitios de la Costa del Pacifico. Ambos representan posibles fronteras, aunque Kaminaljuyu evidentemente estuvo envuelta en algunos acontecimientos con las Tierras Bajas durante el Clásico.
Los estudios de muestras de ADN tomadas de huesos de antiguos Mayas han sido realizados con muy poco éxito (Merriwether et al. 1997), por lo que optó por usar características hereditarias de los esqueletos para examinar diferencias genéticas entre poblaciones. Las dimensiones de los dientes es una característica altamente hereditaria (Kieser 1990; Townsend y Brown 1978), y ha sido usada para examinar relaciones biológicas con más éxito en otras partes del mundo (Brace et al. 1991; Christensen 1998a y b; Powell 1995; Stojanowski 2003, 2004). Las mediciones craneales también suelen usarse en estudios de distanciamiento biológico, aunque no se tomaron en este estudio por cierto número de razones. Primero, los cráneos Mayas, están muy mal conservados. Segundo, la práctica de deformación craneal fomenta la reducción en la utilidad de cráneos de los antiguos Mayas para investigaciones de distanciamiento biológico. Afortunadamente, los dientes de los Mayas de las Tierras Bajas, en general, están bien preservados y presentan poco desgaste, debido a la preferencia de los antiguos Mayas por el maíz procesado en su dieta y por el uso de piedras de moler de piedra caliza, los que no producen mucha arenilla como las de basalto.
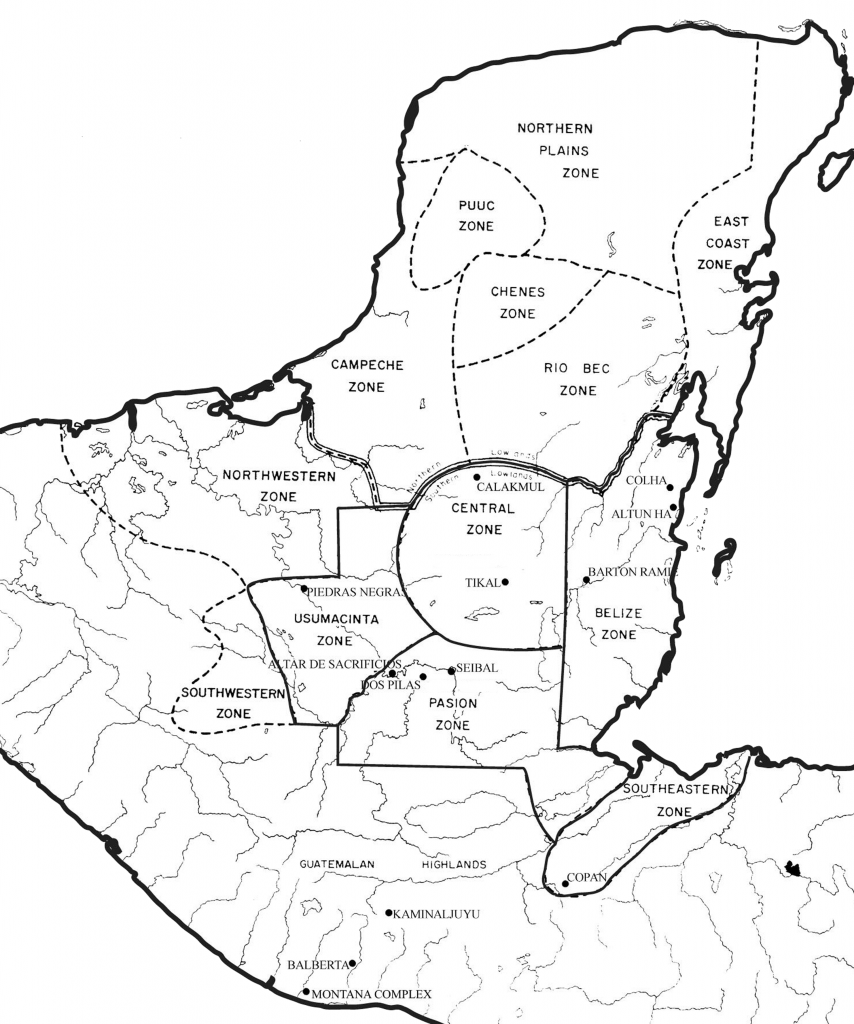
Figura 1 Mapa de los sitios usados en este estudio (modificados de Culbert y Rice 1990:29)
Se usaron medidas mesodistales y bucolinguales en los dientes (Buikstra y Ubelaker 1994). Se excluyeron dientes, incluso con desgaste moderado. Sólo fueron usados los dientes izquierdos en los análisis estadísticos. Si el diente izquierdo estaba ausente, se utilizaba el diente derecho. Antes de cualquier análisis estadístico, se probaron los datos para calcular cualquier fuente de error estándar, incluyendo errores intra-observadores, medidos con una distribución anormal y efectos de la edad. Las medidas que mostraron cualquiera de esas fuentes de error, fueron excluidas. Los datos originales fueron transformados al modo Q, para remover los efectos alométricos y de sexo. Ya que se requieren grupos completos de datos para los análisis de variables múltiples de rasgos cuantitativos, los datos ausentes fueron estimados usando un programa de computación de atribuciones múltiples, llamado NORM. Sin embargo, antes de estimar los datos ausentes, los individuos que habían perdido la mayoría de sus medidas fueron removidos. Después que todos los datos problemáticos fueron removidos, la muestra final se redujo a nueve medidas de 352 esqueletos.
Se usaron tres diferentes métodos estadísticos para examinar la variabilidad de la población del Clásico Maya. Tanto el Análisis de los Principales Componentes, como el Análisis de Distancia de Mahalanobis, dos métodos estadísticos comunes en la antropología física, por lo que no se describirán detalles sobre estos métodos en este momento. La tercera estadística que se usó fue el modelo de la matriz R. Este modelo fue desarrollado por Harpending y Ward (1982), para analizar diferencias en la frecuencia de los alelos entre poblaciones. Relethford y Blangero (1990), modificaron el modelo original de la matriz R, por lo que puede ser aplicado a rasgos cuantitativos de fenotipos, tal es el caso de mediciones dentales. Esencialmente, el modelo de la matriz R trabaja mediante la comparación de heterocigotos genéticos de una muestra de la población, con los heterocigotos de todas las muestras del estudio. Un alto nivel de heterocigoto en una muestra indica un alto nivel de flujo genético. El modelo de la matriz R también puede ser usado para obtener una matriz de distanciamiento genético y la estadística de FST, la cual es una medida de variación entre grupos.
Uno de los beneficios del modelo de la matriz R sobre otras estadísticas en el análisis es que puede moldear los efectos de la deriva genética en la estructura de la población. Las pequeñas poblaciones están sujetas a altos niveles de deriva genética. Cuando el nivel de deriva genética es alto, la frecuencia genética de una población cambiará más rápidamente con cada generación de lo que cambiaría si el nivel de deriva genética fuera pequeño. Entonces, para el control de los efectos de la deriva genética en el modelo de la matriz R, es necesario incorporar datos sobre el tamaño de la población de cada uno de los sitios Mayas. Más que usar estimaciones de población ya publicadas de cada sitio, se utilizó el conteo del número total de estructuras y/o montículos, para determinar el tamaño relativo de la población de cada sitio. Para el análisis de la matriz R, tamaños relativos de población son suficientes, no se necesitan tamaños absolutos de población.
RESULTADOS
El Análisis de los Principales Componentes trabaja reduciendo los datos a una serie de elementos. Los primeros tres del Análisis de los Principales Componentes se muestran aquí (Figura 2). Éstos explican el 67.9% de la variación. Evidentemente, las zonas arqueológicas no producen distintos grupos, como se esperaría con el aislamiento por distancia. Por lo tanto, los sitios de la región del río Pasión, Ceibal y Dos Pilas se agrupan juntos, pero Altar de Sacrificios está aparte. De manera similar, los sitios beliceños de Altun Ha y Colha muestran cierta afinidad en el Análisis de los Principales Componentes. De hecho, Barton Ramie, aparece aislado en la gráfica. Es interesante que los sitios superpotencia de Tikal y Calakmul, ambos localizados en la zona central, caigan en el centro de la gráfica. Yendo más allá, las dos mayores fronteras geográficas y culturales, Kaminaljuyu y la Costa del Pacífico quedan distantes.
El Análisis de Distancia de Mahalanobis fue utilizado para generar una matriz de distanciamiento genético. Pequeños valores en la matriz de distanciamiento genético indican una afinidad biológica cercana. Sin embargo, la matriz de distancia es más bien difícil de manejar y complicada de interpretar. Entonces, se utilizó una escala multidimensional para trazar la matriz de distancia en dos dimensiones (Figura 3). De nuevo, no hay un grupo aparente de sitios por zona arqueológica.
En general, el Análisis de los Principales Componentes y el Análisis de Distancia de Mahalanobis, ilustran resultados similares. Por lo tanto, Kaminaljuyu y la Costa del Pacífico son los límites mayores en ambas gráficas. Además, los sitios beliceños de Altun Ha y Colha se agrupan juntos, incluso Barton Ramie, aparece al límite en ambas gráficas. Aunque también existen algunas diferencias, tal como la posición del sitio de Piedras Negras en la cuenca del río Usumacinta.
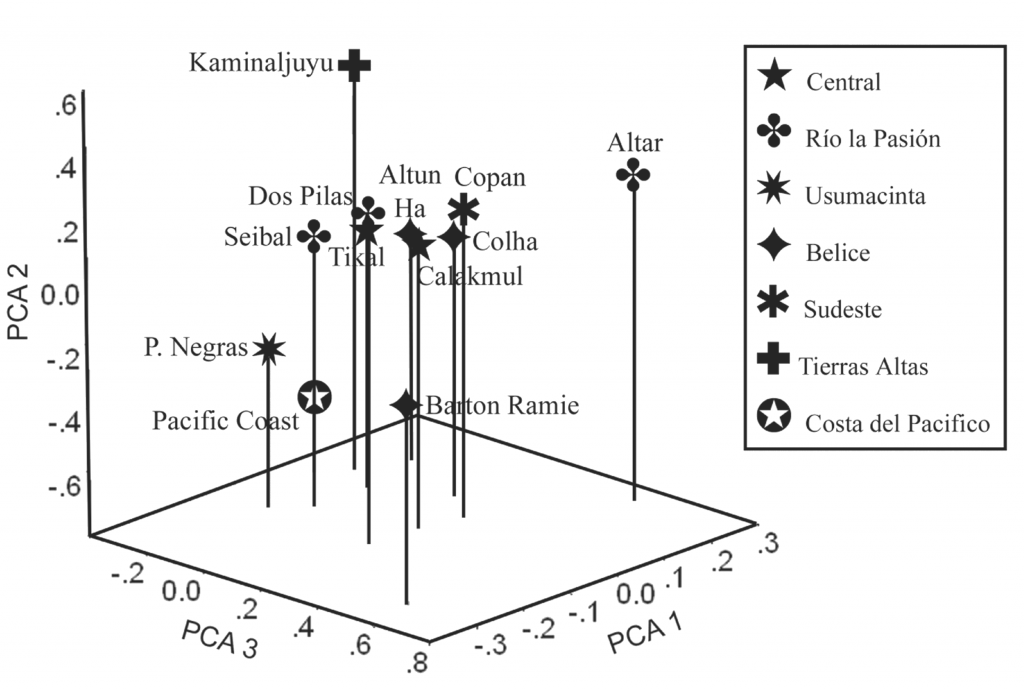
Figura 2 Gráfica de los principales componentes de los sitios del Clásico,
en base a mediciones de rasgos dentales
Volviendo al análisis de la matriz R, se empezará con el cálculo de FST, una medida entre un grupo de variables. Valores bajos para FST indican que los grupos en la muestra presentan una ligera diferenciación genética, mientras que los altos valores para FST indican altos niveles de diferencias biológicas entre los sitios. Se calculó el valor de FST para cada zona arqueológica de las cuales tenia datos de más de un sitio, las que incluyen la zona Central, la del río Pasión y la de Belice (Tabla 1).
En todos los casos, FST fue relativamente baja. Eso resultó especialmente verdadero para la zona central, la cual consiste de los sitios superpotencia de Tikal y Calakmul. Eso es interesante, ya que esos dos sitios fueron acérrimos enemigos a través de gran parte del Clásico. ¿Esos resultados indicarán un fuerte flujo genético entre esos dos sitios? De hecho, el Análisis de los Principales Componentes mostrado con anterioridad, podría darle soporte a tal posibilidad. Sin embargo, seria muy sorprendente si consideramos lo que sabemos sobre el antagonismo entre esos dos sitos.
El distanciamiento biológico se obtuvo por medio del análisis de la matriz R. Una de las fortalezas de la matriz R es que puede ser usada para modelar los posibles efectos de la deriva genética en las dinámicas de antiguas poblaciones. Eso se puede hacer comparando los distanciamientos genéticos que no fueron seriados por el tamaño de la población con aquellas que sí lo fueron. La gráfica que aquí se muestra se basa en los dos principales componentes de una matriz de distanciamiento genético sin escala (Figura 4).
En otras palabras, esta gráfica no toma en consideración la deriva genética. Como se observó con el Análisis de los Principales Componentes y el Análisis de Distancia de Mahalanobis, no se encontró ninguna asociación entre el distanciamiento geográfico y el distanciamiento biológico. Por lo tanto, el patrón general de la gráfica de distancias sin escala es casi siempre similar a aquella que se encontró en los análisis recientes. Cuando se consideran los posibles efectos de la deriva genética, aparecen algunas diferencias generales (Figura 5).
Con más notoriedad, los sitios de Kaminaljuyu, Tikal y Calakmul incrementan su separación de los otros sitios. Estos tres sitios se cuentan entre los tres más grandes en términos del tamaño de la población durante el Clásico. En el caso de Kaminaljuyu, su estatus limítrofe no resulta sorprendente, de hecho es también el más grande límite geográfico y no formó parte directamente de la esfera cultural de las Tierras Bajas Mayas durante el Clásico. El incremento de la separación entre Tikal y Calakmul resultó más sorprendente. Como se mencionó en análisis previos, estos sitios parecen demostrar cierto grado de afinidad y se centran en el medio de la gráfica.

Figura 3 Gráfica de escalas de diferentes dimensiones del Clásico, según las distancias de Mahalanobis
TABLA 1
VALORES PARA FST DE VARIOS DE LOS GRUPOS DE SITIOS EN EL ESTUDIO
Grupos
Numero de Sitios
FST
Error Estándar
Zona Central
2
0.0031
0.0051
Zona del río Pasión
3
0.0181
0.0125
Zona de Belice
3
0.0189
0.0141
Tierras Bajas
10
0.0117
0.0054
Total del Área Maya
12
0.0182
0.0079

Figura 4 Gráfica de los principales componentes de los sitios del Clásico Maya,
en base a la matriz de R sin escala

Figura 5 Gráfica de los principales componentes de los sitios del Clásico Maya,
en base a la matriz de R con escala
¿Y qué es lo que explica la separación que aquí se muestra? Aunque resulta imposible mostrarlo en una figura, pero cuando se examinaron las matrices de distanciamiento genético con y sin escala de donde provienen estas gráficas, emergió una explicación. Cuando la matriz de distanciamiento genético fue seriada por la deriva genética, se incrementó el distanciamiento genético entre Tikal y Calakmul, mientras que sus respectivos distanciamientos con otros sitios del estudio quedaban inamovibles. En otras palabras, cuando tomamos en consideración tamaños de población, la relación entre Tikal y Calakmul se debilita, aunque su relación con otros sitios en la muestra, no cambia.
Con el propósito de entender estos hallazgos, se comparará con los registros arqueológicos y epigráficos. Se sabe que tanto Tikal como Calakmul eran de los más grandes centros del Clásico, con redes de comercio e interacciones políticas que se extendían a través de las Tierras Bajas Mayas. Entonces, la evidencia de flujo genético entre las superpotencias y los otros sitios en el estudio encaja bien con lo que se conoce a partir de los registros arqueológicos y epigráficos. Aparentemente, estas superpotencias no sólo estuvieron intercambiando ideas y mercancías con sitios más pequeños de las Tierras Bajas Mayas, sino también estuvieron intercambiando gente. Eso explicaría la heterogeneidad observada tanto en Tikal como en Calakmul. Pero, la distancia biológica entre Tikal y Calakmul no es pequeña. Entonces, Tikal y Calakmul no interactuaban directamente la una con la otra, más bien, indirectamente a través de otros sitios Mayas. Como indican estos datos, hay evidencia de interacción biológica entre las superpotencias y otros sitios pequeños, pero no hay mucha evidencia de una interacción directamente entre Tikal y Calakmul.
Sin embargo, es bastante probable que determinados sitios interactuaran con Tikal o con Calakmul, dependiendo cual de los dos era particularmente próspero en algún tiempo dado. Eso podría explicar una observación que recientemente hicieran Rice y Forsyth (2004), en consideración con la cerámica de Tikal y Calakmul. Ellos notaron que a pesar de la mutua hostilidad entre Tikal y Calakmul, a través del Clásico ambos tenían estilos cerámicos relativamente similares.
Los resultados de este estudio dan soporte a otro trabajo reciente sobre distanciamiento biológico y de los isótopos estables. En un estudio similar de mediciones dentales se definió que Calakmul mostraba fuertes afinidades biológicas con sitios en el sureste del Petén (Cucina y Tiesler 2004). Este hallazgo destaca la evidencia del flujo genético entre Calakmul y otros sitios de las Tierras Bajas durante el Clásico. De hecho, usando isótopos estables se ha documentado la presencia de numerosos inmigrantes de Petén en tumbas del Clásico Temprano en Kaminaljuyu (Valdés y Wright 2004). De manera similar, Buikstra y sus colegas (2004), han demostrado que los huesos que se cree pertenecieron al fundador de la dinastía de Copan, Yax K’uk’ Mo’, tienen una marca isotópica consistente con el centro de Petén. Cuando los datos genéticos e isotópicos se toman en conjunto, claramente indican que durante el Clásico la población Maya se estaba moviendo, sobreviniendo un flujo genético a través de gran parte de las Tierras Bajas y aún más allá.
CONCLUSIONES
Si se retoma la hipótesis original de investigación: ¿Caracterizó el aislamiento por distancia la estructura de la población del Clásico Maya?, evidentemente, la hipótesis puede ser descartada. Hubo agrupamientos pequeños o no regionales en cualquiera de las gráficas estadísticas, como se hubiera esperado si el aislamiento por distancia explicara la estructura de la población del Clásico Maya. Más bien, la evidencia biológica indica un alto grado de flujo genético entre los sitios Mayas a través del Clásico. Los datos biológicos presentados pueden ser utilizados para explorar muchas otras cuestiones de flujo e intercambio genético entre sitios Mayas. Finalmente, se espera que esta presentación ilustre la importancia de los restos óseos procedentes de los sitios Mayas, como una fuente de información en cuanto a las dinámicas de población de los antiguos Mayas.
AGRADECIMIENTOS
Gracias a mis asesores: Lori Wright, D. Gentry Steele, David Carlson y Rodney Honeycutt. Los permisos para estudiar los restos humanos son del IDAEH, Stephen Houston y Héctor Escobedo (Proyecto Piedras Negras); Frederick Bove (Universidad del Valle); Sharon Misdea y Chris Jones (University Museum, University of Pennsylvania); Herman Helmuth (Trent University); Harry Shafer (Texas A&M University); Tom Hester y Laura Nightengale (Texas Archaeological Research Laboratory); Michelle Morgan y Kristin Bjork (Peabody Museum, Harvard); Vera Tiesler y Andrea Cucina (Universidad Autónoma de Yucatán); y William Folan y Mario Coyoc (Universidad Autónoma de Campeche). Los fondos fueron otorgados por Wenner-Gren Foundation Dissertation Fieldwork Grant (Gr. 7014) y NSF Dissertation Improvement Grant (BCS-0234006).
REFERENCIAS
Brace, C. Loring, S. L. Smith y K. David Hunt
1991 What Big Teeth You Had Grandma! Human Tooth Size, Past and Present. En Advances in Dental Anthropology (editado por M. A. Kelley y C. S. Larsen), pp. 33-57. New York.
Buikstra, Jane. E., T. Douglas Price, Lori E. Wright y James H. Burton
2004 Tombs and Burials in the Early Classic Acropolis at Copan. En Understanding Early Classic Copan (editado por E. E. Bell, M. A. Canuto y R. J. Sharer), pp.191-212. University of Pennsylvania Museum of Archaeology and Anthropology, Philadelphia.
Buikstra, Jane E. y Douglas H. Ubelaker
1994 Standards for Data Collection from Human Skeletal Remains. Arkansas Archeological Survey Research Series No. 44. Arkansas Archeological Survey, Fayetteville.
Christensen, Alexander F.
1998a Colonization and Microevolution in Formative Oaxaca, Mexico. World Archaeology 30 (2):262-285.
1998b Odontometric Microevolution in the Valley of Oaxaca, Mexico. Journal of Human Evolution 34:333-360.
Cucina, Andrea y Vera Tiesler
2004 Dental Morphometry and Biological Affinity in Pre-Contact and Contact Maya Populations from the Peninsula of Yucatan. Mexicon 26 (1):14-19. Möckmühl.
Culbert, T. Patrick (ed)
1973 The Classic Maya Collapse. University of New Mexico Press, Albuquerque.
Culbert, T. Patrick, Laura J. Kasokowsky, Robert E. Fry y William A. Haviland
1990 The Population of Tikal, Guatemala. En Precolumbian Population History in the Maya Lowlands, (editado por T.P. Culbert y D. S. Rice), pp. 103-122. University of New Mexico Press, Albuquerque.
Culbert, T. Patrick y Don S. Rice (ed)
1990 Precolumbian Population History in the Maya Lowlands. University of New Mexico Press, Albuquerque.
Fahsen, Frederico.
2003 La Escalinata Número 2 de Dos Pilas, Petén, Los Nuevos Escalones. Informe presentado a Foundation for the Advancement of Mesoamerican Studies, Inc.
Folan, William J., Joyce Marcus, S. Pincemin, Rosario Domínguez, L. A. Fletcher y A. Morales Lopéz
1995 Calakmul, Campeche: New Data from an Ancient Maya Capital in Campeche, Mexico. Latin American Antiquity 6:310-334.
Harpending, Henry C. y R. H. Ward
1982 Chemical Systematics and Human Populations. En Biochemical Aspects of Evolutionary Biology (editado por M. H. Nitecki), pp. 213-256. University of Chicago Press, Chicago.
Kieser, Julius A.
1990 Human Adult Odontometrics: The Study of Variation in Adult Tooth Size. Cambridge University Press, New York.
Martin, Simon y Nikolai Grube
1995 Maya Superstates. Archaeology 48 (6):41-46.
Merriwether, D. Andrew, David M. Reed y R. E. Ferrell
1997 Ancient and Contemporary Mitochondrial DNA Variation in the Maya. En Bones of the Maya: Studies of Ancient Skeletons (editado por S. L. Whittington y D. M. Reed), pp.208-217. Smithsonian Institution Press, Washington, D.C.
Powell, Joseph. F.
1995 Dental Variation and Biological Affinity among Middle Holocene Human Populations in North America. Tesis de Doctorado. Texas A&M University, College Station, Texas.
Relethford, John H. y John Blangero
1990 Detection of Differential Gene Flow from Patterns of Quantitative Variation. Human Biology 62:5-25.
Rice, Prudence M. y Don W. Forsyth
2004 Terminal Classic-Period Lowland Ceramics. En The Terminal Classic in the Maya Lowlands: Collapse, Transitions, and Transformation (editado por A. A. Demarest, P. M. Rice y D. S. Rice), pp.28-59. University Press of Colorado, Boulder.
Stojanowski, Chrisptopher M.
2003 Differential Phenotypic Variability Among the Apalachee Mission Populations of La Florida: A Diachronic Perspective. American Journal of Physical Anthropology 120:352-363.
2004 Population History of Native Groups in Pre-and Postcontact Spanish Florida: Aggregation, Gene Flow, and Genetic Drift on the Southeastern U.S. Atlantic Coast. American Journal of Physical Anthropology 123:316-332.
Townsend, G. C. y T. Brown
1978 Heritability of Permanent Tooth Size. American Journal of Physical Anthropology 49:497-504.
Valdés, Juan Antonio y Lori E. Wright
2004 The Early Classic and its Antecedents at Kaminaljuyu: A Complex Society with Complex Problems. En Understanding Early Classic Copan (editado por E. E. Bell, M. A. Canuto y R. J. Sharer), pp.337-356. University of Pennsylvania Museum of Archaeology and Anthropology, Philadelphia.