

Iglesias, María Josefa, Sara Álvarez López de Rodas, Eva Fernández Domínguez, Eduardo Arroyo-Pardo, Andrés Ciudad Ruiz y Jesús Adánez Pavón.
2003 Estudio de secuencias de DNA Mitocondrial en una muestra Maya de la ciudad de Tikal, Guatemala. En XVI Simposio de Investigaciones Arqueológicas en Guatemala, 2002 (editado por J.P. Laporte, B. Arroyo, H. Escobedo y H. Mejía), pp.451-463. Museo Nacional de Arqueología y Etnología, Guatemala.
40
ESTUDIO DE SECUENCIAS DE DNA MITOCONDRIAL
EN UNA MUESTRA MAYA
DE LA CIUDAD DE TIKAL, GUATEMALA
María Josefa Iglesias
Sara Álvarez López de Rodas
Eva Fernández Domínguez
Eduardo Arroyo-Pardo
Andrés Ciudad Ruiz
Jesús Adánez Pavón
Este trabajo se enmarca en los resultados que se van obteniendo de forma parcial a partir del proyecto “Los Mayas prehispánicos ante el siglo XXI: Aplicación de análisis de ADN mitocondrial al estudio de las clases sociales de la ciudad arqueológica de Tikal, Guatemala”, iniciado en 1999 con 26 muestras procedentes de la ciudad de Tikal. El proyecto general ha sido financiado por el Programa Sectorial de Promoción General del Conocimiento (SGPICYT: PB97-0278), del Ministerio de Educación y Ciencia de España, y la parte correspondiente a los análisis moleculares ha sido parcialmente financiada con el proyecto PR48/01-9837 de la Universidad Complutense de Madrid.
En el XIV Simposio de Investigaciones Arqueológicas de Guatemala, nos centramos en la presentación del proyecto en sí (Iglesias et al. 2001) y en el del año pasado se adelantaron los resultados de los análisis parciales en ADN nuclear (Iglesias et al. 2002). En esta ocasión ampliamos el marco de posibilidades al ponerlo en relación con los orígenes del poblamiento americano, un tema de gran interés y al mismo tiempo plagado de incógnitas.
Debemos partir de la base, anunciada ya en anteriores presentaciones, de las dificultades inherentes a la muestra elegida; y ello tanto por las limitaciones generales de procedencia general, es decir, por el hecho de hallarse en un contexto ecológico tropical, como por la ausencia de patrones validos de extracción – por supuesto según pautas actuales -, al proceder de excavaciones de los años ochenta, cuando el conocimiento de las vías moleculares de investigación apenas estaba entre el “limbo de los justos” y la visión futurista de algunos científicos.
Si bien los objetivos previos del proyecto se centraban en obtener datos aplicables a la puesta en vías de solución de ciertos problemas relacionados con aspectos sociales internos de gentes que vivieron en el Clásico Temprano de Tikal, en esta ocasión – como acabamos de adelantar – ciertos resultados nos llevan a abordar problemas más generales, tales como los de genética de poblaciones en su más amplio consenso.
-
LA PRESENCIA DEL HOMBRE EN AMÉRICA: DE LA INTUICIÓN A LA MANIPULACIÓN
Desde los tempranos tiempos del siglo XVI en que los españoles pusieron su huella en el nuevo continente, siempre existió curiosidad por conocer el origen del hombre americano. De innegable interés es que los padres Acosta y Gregorio García intuyeran de forma muy temprana el primero un origen asiático o europeo único y el segundo un origen múltiple de los pobladores nativos, mientras que en siglos posteriores, y siguiendo vaivenes histórico-filosóficos, los estudiosos y/o curiosos optaron por otras vías, desde las más estrictamente confesionales, como la de las tribus perdidas de Israel – surgidas de una visión bíblica de la historia -, a las más extravagantes de viajes transatlánticos o transpacíficos. Hay que esperar al siglo XVIII para que los nuevos conocimientos geográficos abran vías más racionales.
-
LA CIENCIA AVANZA
En el siglo XIX, ya en el terreno más científico (Coral et al. 1999), los primeros trabajos se dieron a partir de análisis antropométricos de poblaciones nativas de la actualidad, comparaciones lingüísticas y estudios de artefactos y construcciones antiguas. A pesar de ello aún se plantearon hipótesis extravagantes como la que defendía que el lugar de origen, no ya sólo del hombre americano sino de la humanidad, estaba en la región meridional de América (Ameghino), hipótesis enseguida refutada por investigadores posteriores (Hrdlicka, Keith) que, por otra parte, durante mucho tiempo defendieron la idea de una sola migración mongoloide, aún en sucesivas oleadas. Propugnaban una homogeneidad somática tan fuerte que otro antropólogo (Kate), contemporáneo pero menos restrictivo, se preguntaba si «era una cuestión de amor propio o simplemente estaban aplicando la doctrina Monroe – América para los americanos – a la ciencia del hombre» (Comas 1983:569).
Ciertamente muchas teorías quedaron reducidas a meras anécdotas al avanzar cada vez más determinados conocimientos científicos, dejando el estado de la cuestión del poblamiento bajo estos puntos básicos: a) ausencia de autoctonismo americano, b) inmigración preponderante mongoloide; y c) dudas acerca de cuantos tipos humanos contribuyeron al poblamiento. Juan Comas apuntaba ya en su Manual de Antropología Física que:
«No es posible llegar a conclusiones decisivas si no se cuenta con más abundante material informativo, obtenido con técnicas adecuadas. Pero recordando además que encontramos en los amerindios características que no se explican por la simple inmigración de uno o varios contingentes transpacíficos o transatlánticos. Han tenido que actuar forzosamente las mutaciones, la deriva genética y la selección de manera conjunta, porque – insistimos – lo que se encuentra en el amerindio es algo nuevo que no está presente en las poblaciones del Viejo Mundo, ni es derivable del mismo por el solo mestizaje» (Comas 1983:589-587).
-
LA MODERNIDAD ATACA
Así las cosas, cuando se inician los estudios con datos genéticos el panorama debiera comenzar a cambiar de forma considerable, pero lo cierto es que da la impresión de que, cómo vamos a ver, seguimos con un mismo problema de fondo: el de las grandes lagunas en cuanto a la muestra, de manera que ésta sirva de base para establecer un panorama al menos un tanto fiable. Y si bien las expectativas despertadas por la explosión de las vías moleculares pueden estar ayudando a confirmar o desechar los estudios fisiográficos anteriores, también es cierto que sus – aún – fuertes carencias técnicas están contribuyendo a complicar aparentemente el panorama.
Uno de los primeros estudios de ADNmt en poblaciones americanas actuales se realizó en el suroeste de los Estados Unidos (Wallace et al. 1985), y al compararlo con poblaciones europeas, africanas y asiáticas se pudieron detectar haplotipos o marcadores sólo característicos para poblaciones americanas o que aparecían raramente en poblaciones asiáticas, pero no en las de otros continentes. Sus conclusiones fueron que por la vía asiática habían llegado a América una serie de líneas de descendencia o linajes fundadores que dieron origen a otros nuevos al dispersarse por el resto del continente. Si bien este estudio recibió críticas por lo limitado de la muestra que utilizó, también es cierto que fue una buena base en la que fundamentar estudios posteriores.
Y así se llevaron a cabo estudios similares entre los Pima de Arizona, grupos de Centroamérica, Mayas de Yucatán y Ticuna de América del Sur (Schurr et al. 1990). De los resultados obtenidos se propusieron un total de cuatro linajes fundadores para las diferentes poblaciones americanas. En esa misma vía se añadieron nuevas muestras (Torroni et al. 1992) de poblaciones de Alaska, Canadá y Estados Unidos que confirmaron la idea de los ya propuestos cuatro linajes maternos distintos (A, B, C y D) fundadores de las poblaciones americanas, y que guardan relación con una serie de oleadas diferenciadas que pudieron iniciarse entre hace 40,000 y 20,000 años. Sin embargo, datos publicados posteriormente (Merriwether et al. 1994, 1995) inciden – desde otro punto de vista – en la idea de la existencia de una sola oleada migratoria.
En cualquier caso, otras investigaciones realizadas con nuevas muestras (Bianchi 1994; Easton et al. 1996), ya están planteando la existencia de más linajes fundadores que los cuatro originalmente concebidos, pero que al no estar tan uniforme o ampliamente reconocidos a lo largo del continente, como los ya aludidos A, B, C y D, no han adquirido todavía un rango similar. Vamos a analizar brevemente el histograma de los diferentes linajes propuesto por Meriwether et al. (1994) en el que se ve de una forma bastante clara su presencia espacial desde las muestras procedentes de Siberia a las de una zona meridional de Sudamérica.
A grandes rasgos podemos ver cosas tales como que el Linaje A (Figura 1a) está prácticamente ausente en las muestras siberianas salvo entre los Esquimales o Inuits, y algún grupo más (Chukchi y Koryaks); pero en cuanto entramos en América la presencia se hace muy intensa en Alaska, Canadá y Estados Unidos (salvo excepciones como en el caso de los Pima de Arizona); en la limitada muestra mesoamericana (Mayas de Yucatán) y grupos centroamericanos. A partir del norte de Colombia, los porcentajes de aparición se vuelven irregulares y van descendiendo, hasta llegar a una presencia apenas testimonial en grupos del sur de Chile.
El Linaje B (Figura 1b) es inexistente en Siberia y Alaska y bastante limitado con excepciones en Canadá. Aumenta su presencia en Estados Unidos y Centroamérica, se aprecian carencias casi totales en grupos pre y Amazónicos, y comienzan a aumentar alcanzando unos porcentajes de aparición muy elevados entre poblaciones andinas (Aymará) y menores en los grupos chilenos.
El Linaje C (Figura 2a) presenta amplias fluctuaciones con fuerte presencia siberiana, con significativas lagunas entre esquimales americanos (Inuits) y grupos del noroeste de Canadá, alguno del sureste de los Estados Unidos y especialmente en América Central. En América del Sur está presente en norteños colombianos y amazónicos, mientras es irregular entre los grupos andinos y acaba aumentando en los meridionales grupos chilenos.
El Linaje D (Figura 2b) aparece en niveles variables en grupos de Siberia, de Alaska y Canadá, disminuyendo significativamente entre las muestras de Estados Unidos y Centroamérica. En Sudamérica es de interés reseñar los altos porcentajes de aparición a excepción de los grupos andinos (Aymará).
Llegado este momento es necesario indicar que las muestras de ADNmt antiguo contenidas en los estudios de Merriwether et al. (1994, 1995) representan un porcentaje muy pequeño: 58 (16 de momias del norte de Chile – 12 Chinchorro y 4 Incas -, 12 de Virginia Occidental y 30 de Copan) frente a las 3,256 de las muestras actuales, decir, la muestra de grupos modernos supone en su estudio un 98.21%, frente a un 1.78 % de grupos prehispánicos.
Los resultados interrelacionados de prehispánicos y actuales, nos muestran que los individuos de los que proceden los tejidos momificados chilenos están lejos de los grupos andinos actuales y algo más cercanos a grupos «Araucanos» como los Pehuenches, Mapuches o Huilliches. Lo mismo sucede en las muestras de Virginia respecto a los grupos de áreas aledañas norteamericanas.
En el caso de Copan, las afinidades más directas debieran darse teóricamente con las muestras de Mayas actuales de Yucatán, pero sucede lo mismo que en los casos anteriores. Más específicamente, los autores nos informan que han podido asignar a las muestras de Copan pertenencia a los Linajes C o D, mientras que los Yucatecos actuales, aunque aparecen en los cuatro linajes se adscriben fundamentalmente a los Linajes A y B.
Como en el caso del comentario que en párrafos anteriores hacía Juan Comas en los años sesenta, los mismos autores reconocen que sin duda el pequeño tamaño de la muestra puede ser la razón de los – hasta cierto punto – no esperados resultados.
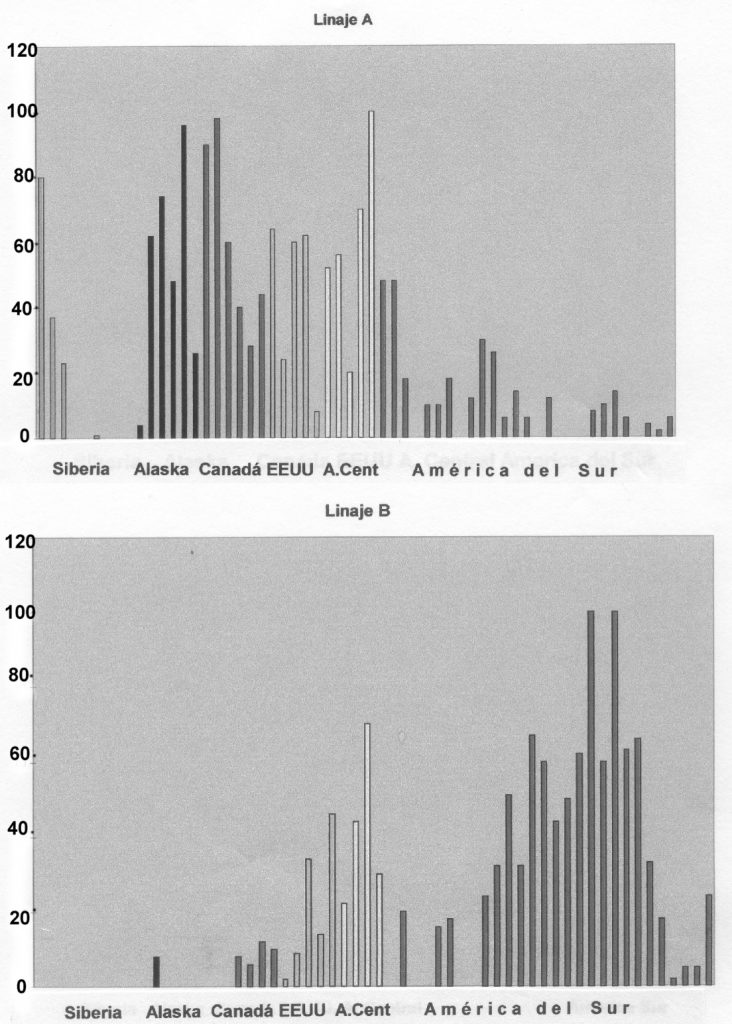
Figura 1 a) Histograma del Linaje A; b) Histograma del Linaje B (Modificados de Merriwether et al. 1994)
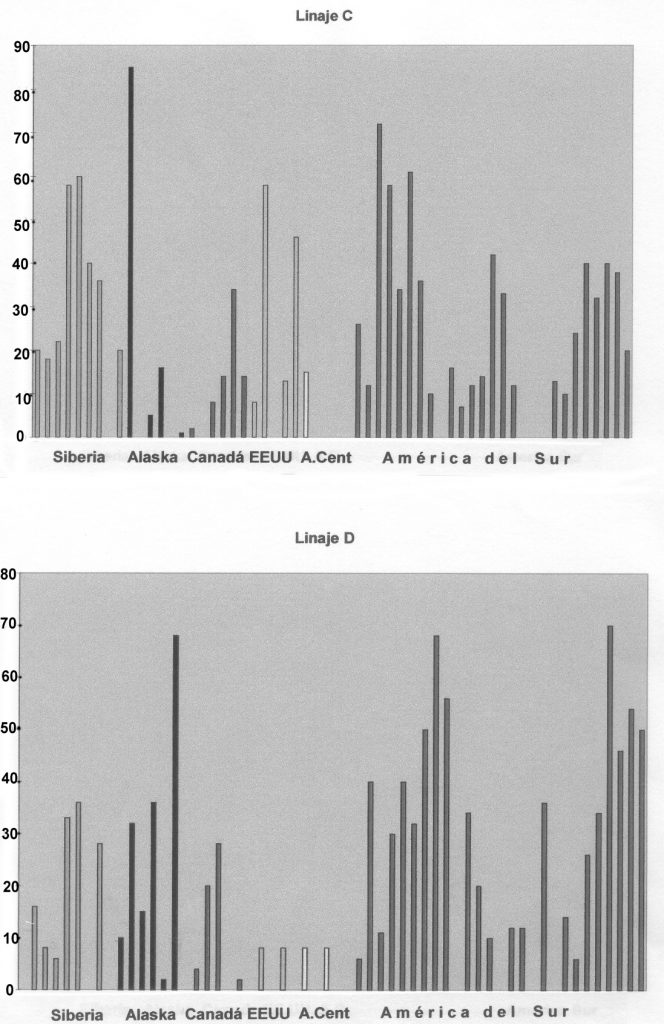
Figura 2 a) Histograma del Linaje C; b) Histograma del Linaje D (Modificados de Merriwether et al. 1994)
-
ALGUNOS RESULTADOS DE LA MUESTRA DE TIKAL
Para esta ocasión se inició el estudio sobre un conjunto de 18 muestras de dientes, de las que sólo seis dieron resultados. La parte correspondiente al análisis molecular ha sido elaborada basándose en el informe «Análisis de DNA mitocondrial en muestras antiguas de la ciudad de Tikal (Guatemala)» realizado por Sara Álvarez López de Rodas y Eduardo Arroyo-Pardo del Departamento de Toxicología y Legislación Sanitaria, en la Facultad de Medicina de la Universidad Complutense de Madrid, y del Eva Fernández Domínguez de la Unitat d’Antropologia del Departamento de Biología Animal en la Facultad de Ciencias Biológicas de la Universidad de Barcelona.
Se procedió a extraer ADN en dos procesos independientes: uno con 11 muestras y otro con los siete restantes. El proceso de extracción de ADN comenzó por limpiar la superficie de los dientes con óxido de aluminio disparado a alta presión, con la ayuda de una Sandblaster Dentalfarm Base-1 Plus. Durante todo el proceso se tomaron todas las medidas necesarias para evitar la contaminación de las muestras con ADN exógeno; esto lleva consigo la limpieza de todos los materiales utilizados con detergente, agua destilada, etanol 70% y radiación con luz UV.
Una vez limpiados, los dientes se trituraron en un molino de nitrógeno líquido, y el polvo obtenido se lavó con una disolución de EDTA 0.5M pH 8.0. Tras centrifugar durante 5 minutos a 2500 rpm se descartó el sobrenadante y se repitió el lavado. Posteriormente el pellet se incubó en una solución de lisis que contenía EDTA 50mM, Tris 100mM y proteinasa K (20 mg/ml) toda la noche a 37ºC con agitación. Se volvieron a centrifugar las muestras a 2500rpm durante 5 minutos y se siguió trabajando con el sobrenadante. Se añadieron 5 ml de fenol/cloroformo/isoamílico y tras agitar durante 15 minutos se centrifugó 2 minutos a 3000 rpm. Se separó la fase acuosa, que se concentra con Centriplus, obteniéndose un volumen final de 100-150l.
Para asegurar que el ADN de las muestras no había sido contaminado durante el proceso, se introdujeron controles de extracción (sólo contienen los reactivos y sigue el proceso de extracción a la vez que las muestras); 2 en la primera ronda de 11 muestras y 1 en la siguiente ronda de 7. En el proceso de amplificación se amplificó junto con las muestras el control de extracción correspondiente.
En cuanto a amplificación, las secuencias de los cebadores corresponden a la región HV-1 del ADNmt y amplifican dos regiones solapadas -denominadas 1 y 2 en la tabla de resultados- según puede verse en Fernández et al. (2000).
Desgraciadamente, excepto para dos de las muestras (TK2 y TK8) la información obtenida es parcial, pues únicamente se ha conseguido secuenciar uno de los dos fragmentos (167bp) de una región o sección de estudio del ADNmt (un segmento de 304bp de la región Hipervariable I del mtDNA [HV-1]). Probablemente esto se debe a que, a pesar de tratar de realizar una selección aparentemente cuidadosa de las piezas dentarias (completas y sin fracturas), en el ámbito molecular las muestras en general están bastante mal preservadas, sin duda debido al clima extremadamente caluroso y húmedo de la región.
Para tener una correcta asignación étnica y de haplogrupo se necesita la información completa de toda la región o sección. Al no ser esto posible por el momento, se ha tratado de hacer una asignación aproximada con la información disponible hasta el momento. Dicha asignación es pues susceptible de cambio y debe ser por tanto observada con cautela.
El estudio preliminar muestra una elevada heterogeneidad dentro de las muestras analizadas. Buscando amplias afinidades con las bases de datos que actualmente se manejan, étnicamente se distinguen hasta tres orígenes para las muestras de Tikal estudiadas: Europeo, representado por las muestras TK9, TK13 y TK6 y TK 2, Europeo-Americano, representado por la muestra TK8, y Africano, representado por la muestra TK3. Así mismo, una de las muestras, TK7, presenta una combinación de mutaciones que no aparece en una amplia base de datos de poblaciones modernas (The Concordance of Nucleotide Substitutions in the Human mtDNA Control Region. University of Cambridge), utilizada para realizar la asignación étnica de las muestras antiguas.
El tipo de controles técnicos efectuado en el laboratorio nos induce a pensar que ha sido un proceso plenamente controlado y que en ningún caso hay contaminación en el ámbito de extracción, al no concordar ninguna muestra con la investigadora que realizó el trabajo.
En cualquier caso, hay que tener en cuenta que incluso los linajes europeos hallados en la muestra se encuentran en Europa con una probabilidad bastante baja y, aunque lo más fácil es pensar en una contaminación con ADN de las personas relacionadas con el trabajo de campo arqueológico (arqueólogos, excavadores), hay que recordar que la genética de poblaciones del pasado pudiera ser diferente a la actual, y que en buena medida algunos resultados nos llevan por ese camino.
La presencia de un posible linaje africano entre las muestras puede explicarse de la misma manera, dada la heterogeneidad actual de las poblaciones de Petén y su ya antigua relación con población beliceña de origen africano, que puede estar presente en un cierto porcentaje de los excavadores que intervinieron en los diferentes procesos arqueológicos. Por motivos obvios no se contempla la lejana idea de un linaje ancestral procedente de rutas transatlánticas africanas, y que entraría en patente contradicción con todas las teorías actuales formuladas hasta el momento para la genética de poblaciones antiguas y modernas.
Lo más sorprendente de los aparentemente decepcionantes resultados, es la ausencia en la muestra de Tikal de haplogrupos típicamente americanos (A, B, C, D) que Merriwether et al. (1994) encontraron en sus muestras de Copan, ya que incluso pensando que estamos ante unos resultados contaminados, existe un sustrato americano indudable en un porcentaje elevado de las personas implicadas en las investigaciones de los años 80-90 y por ello cabe pensar que debiera aparecer alguno de los linajes americanos propuestos (A, B, C y D).
-
LA MUESTRA TK8
Diferente es el caso de una de las muestras (TK8) que pertenece al haplogrupo X, variante mitocondrial presente en un muy bajo porcentaje en poblaciones europeas (y ello excluiría en gran medida la contaminación), y que en cambio ha sido encontrada (Figura 3) en una frecuencia considerable en ciertas poblaciones nativas norteamericanas (25% Ojibwa, 15% Sioux, 11-13% Nuu-Chah-Nulth, 7% Navajo, 5% Yakima; Torroni et al. 1993; Brown et al. 1998, Scozzari et al. 1997).
Desde su descubrimiento, ciertos autores han propuesto que el haplogrupo X, junto con los ya descritos A, B, C y D, puede constituir el quinto linaje fundador de las poblaciones americanas (Ward et al. 1991, 1993; Bailliet et al. 1994; Forster et al. 1996). A diferencia de los otros haplogrupos típicamente americanos (A, B, C, D) el haplogrupo X parece estar ausente en poblaciones asiáticas.
En su estudio de 1998, Brown et al., a partir de los valores de afinidad de las variantes europea y americana de dicho haplogrupo X, proponen que su penetración en América se produjo desde poblaciones europeas de Oriente Medio hace 23,000-36,000 años, o bien hace 12,000-17,000 años. En sus investigaciones, dichos autores descartan la mezcla reciente con poblaciones europeas y americanas como explicación de la presencia del haplogrupo X en estas últimas, basándose en la ausencia de otras variantes europeas en América, en las diferencias existentes entre los haplotipos europeos y los americanos y en la divergencia encontrada en las variantes americanas, signo de una llegada antigua de estas líneas al Nuevo Continente.
Apoyando esta idea, existen evidencias de la presencia de dicho haplogrupo X en muestras de la época precolombina, tales como las secuencias parciales recuperadas por Hauswirth et al. 1994, Ribeiro-Dos-Santos et al.1996, portadoras de las mutaciones 16223T y 16278T. Stone y Stoneking en sus trabajos de 1993 y 1998 con población oneota de ESTADOS UNIDOS. De 1300 DC, encontraron dos muestras del total de 52 analizadas que no pertenecían a los haplogrupos A, B, C y D. Estas muestras compartían los motivos 16223T, 16278T (además de 16093T, 16093C, 16189C, 16227G y 16357C) y pueden por lo tanto asignarse al haplogrupo X. Nuestro análisis, por lo tanto, estaría en esta línea, apoyando la hipótesis de un nuevo linaje americano propuesta por estos y otros autores.

Figura 3 Histograma del Linaje X (Modificado de Merriwether et al. 1994)
CONTEXTO ARQUEOLÓGICO DE LA MUESTRA TK8
La muestra TK8 que ha aportado estos resultados corresponde al Entierro 177 procedente del Grupo 6D-V de Tikal (Figura 4), y que hemos fechado para finales el Clásico Temprano. Esta inhumación se halló al realizar un pozo en el interior de una plataforma localizada en el lado sur del asentamiento. El enterramiento se encuentra colocado en un chultun de 1.30 m de altura y 1.10 m de anchura máxima, tapiado en su cara este y que a su vez está cubierto o «señalado» por una tosca construcción en cuatro niveles que hemos denominado «altar» (1.35 m norte-sur, 1.30 m este-oeste y 0.94 m de altura máxima).
Lo más interesante de esta inhumación (Figura 5a), es que los restos humanos se hallaron depositados en un inhabitual continente: una gran vasija de base cónico-truncada y paredes rectas que alcanzan una altura de 0.70 m; el borde está reforzado y su diámetro es de 0.68 m (Iglesias 1987; Laporte et al. 1992; Figura 5b).

Figura 4 Localización del Grupo 6D-V de Tikal

Figura 5 Entierro 177: a) Planta; b) Perfil
La inhumación es primaria, indirecta e individual, de un adulto medio masculino (Fialko 1987; Wright 1996). Aparentemente se conservan todos los huesos, aunque muchos de ellos aparecen fracturados, incluido el cráneo. Dos dientes presentan características especiales: uno de ellos una mutilación del tipo B-5 y un segundo una incrustación circular posiblemente de pirita. También en el interior, presumiblemente sirviendo de collar al muerto, se hallaron tres cuentas de jade con perforación bicónica, y dos conchas del género spondylus con perforación en la parte superior. En torno a la vasija contenedora, se colocaron además otras siete piezas cerámicas (dos platos, tres cilindros de patas-almena y dos cuencos).
Sin duda se trata de un enterramiento excepcional por lo infrecuente de sus características formales, al ser el único contenido en una gran olla aparecido no sólo en Tikal, sino en otras ciudades Mayas de las Tierras Bajas. El conjunto nos indica un complejo funerario de élite. Sin que vayamos a realizar ningún tipo de especulación si es interesante reseñar que curiosamente esta muestra aúna un resultado sorprendente en el ámbito molecular con un contexto cultural hasta cierto punto excepcional.
CONSIDERACIONES FINALES
Como resumen de esta presentación hay que concluir que, por una parte, el hallazgo en una de las muestras de las mutaciones 16223T y 16278T, ambas definitorias del haplogrupo X, vendría a apoyar la hipótesis propuesta por Brown et al. (1998) y otros autores sobre una quinta línea de descendencia o linaje fundador, y supone un dato de enorme interés para el estudio del poblamiento de América, ya que los haplogrupos X encontrados hasta la fecha eran originarios de regiones más al norte. Sin embargo, sería importante estudiar más profusamente las muestras hasta poder secuenciar, si es que ello es posible, el segundo fragmento de las muestras restantes.
Los resultados del resto de las muestras, aunque pendientes de ser corroborados, apuntarían más bien a una contaminación actual debida a la lógica falta de precauciones en las sucesivas manipulaciones en el contexto arqueológico que hacia una hipótesis, hasta el momento sin precedentes, de la existencia en América de variantes mitocondriales europeas actualmente de baja frecuencia.
Teniendo en cuenta la naturaleza de los datos y la escasa información disponible, cabe ser cautelosos y excluir primero la contaminación exógena como explicación de los resultados obtenidos. Para ello sería indispensable y de gran interés científico la secuenciación del fragmento faltante en estas muestras, así como la caracterización genética de los miembros del laboratorio que han intervenido directamente en el análisis de las muestras y, especialmente, de los arqueólogos y excavadores responsables de la investigación de campo, aspecto éste difícil de conseguir.
Sin duda la degradación y/o contaminación de las muestras son dos de los grandes problemas con que nos encontramos actualmente. Por ello se hace imprescindible una mayor investigación en el campo de la biología molecular, de manera que se pueda hacer uso fiable de los miles y miles de restos humanos excavados en toda América a lo largo de muchos años. Es imposible renunciar a todo este bagaje y limitar la reconstrucción del desarrollo poblacional de América a partir de muestras tomadas en aquellos lugares en los que los dioses nos sean extremadamente favorables, aquellos yacimientos arqueológicos en los que confluyan una ecología amable que nos preserve extraordinariamente los restos humanos y una asepsia en el proceso de excavación tan parecida a una intervención quirúrgica como imposible en el trabajo real de campo.
Por último nos gustaría realizar una reflexión final en torno a los estudios de genética de las poblaciones americanas. Hasta el momento, las bases de datos que se han manejado para definir los cuatro o cinco linajes fundadores de la población americana se han centrado en la toma de muestras en grupos nativos actuales. Aun cuando pensáramos –y es bastante improbable- que en estos grupos no ha existido ningún tipo de mestizaje a lo largo de los siglos de interacción con poblaciones del resto de los continentes (Europa, África y Asia), llama la atención que no muestren un grado de afinidad mayor con las muestras de poblaciones antiguas (Chile, Estados Unidos y Honduras).
Es comprensible que resulte más atractiva la resolución de problemas tales como el origen del hombre americano que la dinámica de cambios genéticos que hayan podido originarse a raíz de la invasión de europeos en el siglo XVI; europeos que por otra parte – al menos en el caso de los españoles que en última estancia fueron mayoritarios en una buena parte del continente – conllevaban una mezcla genética ya de por sí importante, que puede rastrearse a nivel histórico-cultural desde al menos el Neolítico, con entradas de pueblos procedentes de la Europa Central, el Mediterráneo en un sentido genérico (Grecia, Roma, Fenicia, Cartago, etc), y las más tardías e importantes intrusiones árabes del siglo VIII de nuestra era que afectaron prácticamente a toda la península Ibérica hasta la etapa de la conquista de América, un total de 700 años de mestizaje.
Se hace evidente que, junto a una mejora de las técnicas de análisis molecular – que sin duda llegará poco a poco -, es necesario aumentar la muestra lo más controlada posible de poblaciones americanas antiguas, y al tiempo trabajar con poblaciones surgidas a lo largo de los más de 500 años de mestizaje, de manera que aumentando el número de árboles la visión del bosque humano sea cada vez más comprensible para todos nosotros.
-
REFERENCIAS
Bailliet G., F. Rothhammer, F.R. Carnese, C.M. Bravi y N.O. Bianchi
1994 Founder Mitochondrial Haplotypes in Amerindian Populations». American Journal of Human Genetics 55:27-33.
Brown M.D., S.H. Hosseini, A. Torroni, H.J. Bandelt, J.C. Allen, T.G. Schurr, R. Scozzari, F. Cruciani y D.C. Wallace
1998 mtDNA Haplogroup X: An Ancient Link Between Europe/Western Asia and North America. American Journal of Human Genetics 63:1852-1861.
Comas, Juan
1983 Manual de Antropología Física. Instituto de Investigaciones Antropológicas. UNAM, México.
Easton, R.D., D.A. Merriwether, D.E. Crews y F.E. Ferrell
1996 MtDNA Variation in the Yanomami: Evidence for Additional New World Founding Lineages. American Journal of Human Genetics 59:213-225.
Fernández, E., J. García-Bour, I. Arenal, A. Pérez-Pérez, L. Valdés y D. Turbón
2000 Hacia el origen de los vascos. Secuencias de ADM mitocondrial antiguo del País Vasco. Complutun 11:101-111.
Fialko, Vilma
1987 Informe sobre el análisis osteológico de los restos del Proyecto Nacional Tikal. Informe, Proyecto Nacional Tikal, IDAEH y Escuela de Historia, USAC, Guatemala.
Forster P., R. Harding, A. Torroni y H.J. Bandel
1996 Origin and Evolution of Native American mtDNA Variation: A Reappraisal. American Journal of Human Genetics 59:935-945.
Hauswirth, W.W., C.D. Dickel y D.A. Lawlor
1994 DNA Analysis of the Windover Population. En Ancient DNA (editado por B. Herrmann y S. Hummel), pp.104-121. Springer, Nueva York.
Iglesias Ponce de León, María Josefa
1987 Excavaciones en el Grupo Habitacional 6D-V, Tikal, Guatemala. Tesis Doctoral, Universidad Complutense, Madrid.
Iglesias, María Josefa, Andrés Ciudad, Eduardo Arroyo, Jesús Adánez y Sara Álvarez
2001 Aplicaciones de la Antropología Molecular a la Arqueología Maya: El caso de Tikal. En XIV Simposio de Investigaciones Arqueológicas en Guatemala, 2000 (editado por J.P. Laporte, H. Escobedo, A.C. de Suasnávar y B. Arroyo), pp.1069-1088. Museo Nacional de Arqueología y Etnología, Guatemala.
Iglesias, María Josefa, Eduardo Arroyo, Andrés Ciudad, Sara Álvarez, Eva Fernández y Jesús Adánez
2002 Los análisis de ADN de enterramientos de Tikal: la arqueología entre la esperanza y la desesperación. En XV Simposio de Investigaciones Arqueológicas en Guatemala, 2001 (editado por J.P. Laporte y B. Arroyo), pp.875-885. Museo Nacional de Arqueología y Etnología, Guatemala.
Laporte, Juan Pedro, Bernard Hermes, Lilian Vega de Zea y María Josefa Iglesias
1992 Nuevos entierros y escondites de Tikal. Subfases Manik 3a y 3b. Cerámica de Cultura Maya 16:30-68.
Merriwether, D.A., F. Rothhammer y R.E. Ferrell
1994 Genetic Variation in the New World: Ancient Teeth, Bone, and Tissue as Sources of DNA. Experientia 50:592-601.
1995 Distribution of the Four Founding Lineage Haplotypes in Native Americans Suggests a Single Wave of Migration for the New World. American Journal of Physical Anthropolgy 96:411-428.
Merriwether, D.A., W.W. Hall, A. Vahlne y R.E. Ferrell
1996 MtDNA Variation Indicates Mongolian May Have Been the Source for the Founding Population for the New World. American Journal of Human Genetics 59:204-212.
Ribeiro dos Santos, A.K.C, S.E.B. Santos, A.L. Machado, V. Guapindaia y M.A. Zago
1996 Heterogeneity of Mitochondrial DNA Haplotypes in Pre-Columbian Natives of the Amazon Region. American Journal of Physical Anthropology 101:29-37.
Scozzari R., F. Cruciani, P. Santolamazza, D. Sellitto, D.E. Cole, L.A. Rubin, D. Labuda, et al.
1997 MtDNA and Y Chromosome-Specific Polymorfisms in Modern Ojibwa: Implications About the Origin of Their Gene Pool. American Journal of Human Genetics 60:241-244.
Schurr, T.G., S.W. Ballinger, Y.Y. Gan, J.A. Hodge, et al.
1990 American Mitochondrial DNAs Have Rare Asian Mutations at High Frequencies Suggesting They Derived From Four Primary Maternal Lineages. American Journal of Human Genetics 46:613-623.
Stone, A.C. y M. Stoneking
1993 Ancient DNA From a Pre-Columbian Amerindian Population. American Journal of Physical Anthropology 92:463-471.
1998 MtDNA Analysis of a Prehistoric Oneota Population: Implications for the Peopling of the New World. American Journal of Human Genetics 62:1153-1170.
Torroni, A., T.G. Schurr, M.F. Cabell, M.D. Brown, J.V. Neel, M. Larsen, D.G. Smith, et al.
1993 Asian Affinities and Continental Radiation of the Four Founding Native American mtDNAs. American Journal of Human Genetics 53:563-590.
Wallace, D.C., K. Garrison y W.C. Knowler
1985 Dramatic Founder Effects in Amerindian Mitochondrial DNAs. American Journal of Physical Anthropology 68:149-155.
Ward, R. H., B. L. Frazier, K. Dew-Jager, S. Pääbo
1991 Extensive Mitochondrial Diversity Within a Single Amerindian Tribe. Proceedings of the National Academy of Science USA 88:8720-8724.
Wright, Lori E.
1996 The Inhabitants of Tikal: A Bioarchaeological Pilot Project. Reporte Final, Foundation for the Advancement of Mesoamerican Studies/Texas A & M University, College Station.