

Descargar este artículo en formato PDF
Wright, Lori E.
1993 La dieta antigua en la región del río de La Pasión. En VI Simposio de Investigaciones Arqueológicas en Guatemala, 1992 (editado por J.P. Laporte, H. Escobedo y S. Villagrán de Brady), pp.172-179. Museo Nacional de Arqueología y Etnología, Guatemala.
14
LA DIETA ANTIGUA EN LA REGIÓN DEL RÍO DE LA PASIÓN
Lori E. Wright
Investigaciones de los sistemas de producción económica han sido un tema común en la arqueología del área Maya. El objetivo principal ha sido la documentación de las estrategias agronómicas empleadas por esta compleja civilización en el severo ambiente de las Tierras Bajas. Pero el estudio de la composición de la dieta antigua ha sido un tema más esquivo. Estudios de restos paleobotánicas han ilustrado la variedad de alimentos explotados por los Mayas. Además de los productos agrícolas importantes como maíz, frijol, pepitoria y chile, estas investigaciones han identificado muchas plantas silvestres que podrían haber sido usadas, como el ramón (Puleston 1982), zapote, nance y una variedad de palmas (Lentz 1991). La identificación de huesos de animales encontrados en excavaciones da un idea de las especies de animales explotadas en la antigüedad, incluyendo una variedad de animales terrestres y acuáticas (Pohl 1985). Desafortunadamente, el pequeño tamaño de las muestras excavadas de restos alimenticios en las Tierras Bajas húmedas limita la interpretación de estos datos.
Sin embargo, nuevos métodos químicos refinados durante los últimos 15 años ofrecen una oportunidad para investigar la dieta en más detalle. El análisis de isótopos estables de carbono y nitrógeno en huesos arqueológicos da una nueva visión de la dieta prehistórica, permite la comparación dietética entre poblaciones antiguas al nivel del individuo. Por ejemplo, por medio de datos químicos se puede estudiar patrones de consumo en poblaciones estratificadas. Además, nos da una oportunidad de investigar cambios en la dieta prehispánica, en relación con el desarrollo de la civilización antigua. En el área Maya, teorías agrícolas del origen y colapso cultural son comunes, pero ha sido difícil verificarlas con los métodos arqueológicos tradicionales.
El análisis isotópico se basa en el proceso del metabolismo de los alimentos en el cuerpo, que traslada la marca química de la dieta al hueso del consumidor. La proporción relativa del isótopo carbono 13 al 12, δ13C (calibrado en relación al estándar de «Pee Dee Belemnite»), refleja la base metabólica de las plantas consumidas. El isótopo carbono 13 es más abundante en plantas que producen un producto compuesto de 4 átomos de carbono llamado C4, que en plantas del tipo C3. Por lo tanto, el consumo de plantas C4 se presenta con un nivel alto de δ13C en el colágeno de huesos, típicamente cerca de 27%o, isotópicamente pesado. En contraste, plantas C3 muestran δ13C de 12%o, o son isotópicamente ligeras (Schwarcz y Schoeninger 1991; van der Merwe 1982). En el área Maya, el consumo del maíz (Zea mays) se identifica por un nivel pesado del δ13C en hueso, porque el maíz es casi la única planta del C4 en el área. Las demás plantas cultivadas y silvestres son del tipo C3 y muestran niveles ligeros del δ13C.
El nivel de los isótopos estables de nitrógeno, δ15N, refleja los procesos de fijación de nitrógeno por plantas. Por ejemplo, plantas leguminosas, como frijoles, muestran niveles bajos del isótopo nitrógeno 15 con respecto al isótopo 14 (cerca de 0%o) que en plantas noleguminosas (DeNiro 1987; Schwarcz y Schoeninger 1991). También, isótopos de nitrógeno ayudan a diferenciar entre fuentes de proteína, en la que la fauna acuática se identifica con niveles más pesada que los de la fauna terrestre (Katzenberg 1989).
Aquí se reportan los resultados preliminares del análisis isotópico de 101 muestras de huesos humanos arqueológicos de la región del río Pasión, en las Tierras Bajas Mayas. Los entierros provienen del periodo Preclásico hasta el Clásico Terminal, de los sitios de Altar de Sacrificios y Ceibal; y del Clásico Tardío en Dos Pilas, Aguateca e Itzan. La mayoría de estos sitios experimentaron el colapso de la cultura elitista al final del periodo Clásico Tardío. En Dos Pilas y Aguateca, el colapso fue abrupto y probablemente estuvo relacionado con conflicto (Demarest 1990; Inomata et al 1991). En Altar de Sacrificios, el proceso parece haber sido gradual, con una ocupación sustancial que continuó después de que el sitio dejó de ser un centro importante de poder político. En contraste, Ceibal mantuvo autoridad política en el periodo Clásico Terminal con aumento de población y nuevas tradiciones arquitectónicas (Mathews y Willey 1991; Willey 1973, 1990).
Frecuentemente, hipótesis agrícolas han sido propuestas para entender el crecimiento y la caída de los sitios Mayas en la región del río Pasión. Por ejemplo, Sharer (1977) propuso que el limitado acceso a tierras agrícolas extensivas creó competencia entre los grupos políticos y que ésta produjo el colapso. En contraste, Adams (1983) propuso que el intenso cultivo de chinampas produjo un excedente que fue el foco del conflicto. Recientemente, Demarest (1990) ha hipotetizado que la guerra expansionista produjo una contracción de los sistemas agrícolas y un colapso ecológico localizado. Esperamos que el análisis isotópico pueda ofrecer nueva información acerca de la naturaleza y evolución de las estrategias dietéticas en la región.
MÉTODOS
Muestras de hueso se seleccionaron de cada entierro adulto con buena preservación del hueso. La mayoría de muestras se tomaron del fémur, pero se tomaron de otros huesos largos en algunos pocos esqueletos que no presentaron fémur. Además, se analizaron cinco muestras de venado (Odocoileus virginianus y Mazama americana) arqueológicos excavados por el Proyecto Petexbatun y vértebras de cinco peces modernos (blanco, Petenia splendida; colorada y guapote, Ciclasoma sp.) de la laguna Petexbatun, para obtener datos básicos sobre fuentes de variación dietética en el ecosistema.
Para la investigación de isótopos estables, las muestras fueron analizadas en el laboratorio de la Doctora Margaret Schoeninger de la Universidad de WisconsinMadison. La muestra de hueso fue limpiada, descalcificada con 0.5 M EDTA a 4C y purificada con 0.125 M hidróxido sódico (NaOH) para eliminar la contaminación de los ácidos húmicos. El colágeno recogido fue secado en frío, calentado con óxido cúprico (CuO) a 850C por una hora y los vapores producidos fueron analizados por espectrometría de masas.
El colágeno fue recolectado en un buen estado de preservación de casi todas las 101 muestras preparadas. Solamente seis muestras no produjeron un seudomorfo bueno de colágeno. Para la mayoría de las muestras se calculó una razón de carbono a nitrógeno (C/N) entre 3.0 y 3.5, indicando buena composición química del colágeno colectado (DeNiro 1985). Los datos isotópicos de solamente cuatro muestras fueron descartados porque los resultados estuvieron fuera de este rango, dando un total de 91 muestras con buenos datos. Diferencias en composición isotópica se identificaron con el análisis de varianza, con una probabilidad de 0.05.
RESULTADOS
El análisis de los huesos de fauna ilustra diferencias en la composición isotópica entre la proteína terrestre y acuática en el ecosistema del río Pasión. El hueso de venado presenta δ13C entre 14.0 y 21.6 %o, una variación ancha. El nivel alto de δ13C en dos de estos animales podría sugerir que los venados invadían las milpas para comer el maíz de vez en cuando y que ellos no solo consumieron plantas salvajes del tipo C3, un patrón que también es encontrado en otras partes (Price et al 1985). Los peces muestran niveles muy ligeros en δ13C, entre 28.6 y 30.8%o. Con respecto al nitrógeno, los venados se caracterizan entre 2.2 y 6.6%o, similar a las plantas noleguminosas. Los peces son más pesados en δ15N, entre 9.2 y 12.3%o, una diferencia que podría ayudar a identificar el consumo de proteína acuática (Katzenberg 1989).
En general, los huesos humanos de la región del río Pasión se caracterizan por niveles de δ13C entre 13.5 y 6.5 %o, lo que indica una dieta basada en el maíz, aumentada con plantas silvestres o cultivadas del tipo metabólico C3. Los resultados de δ15N se calcularon entre 6.5 y 11.4%o, se indica que el consumo tuvo una mezcla de fuentes de proteína. La distribución ancha de estos datos indica una variación considerable en la composición de la dieta entre miembros de la población.
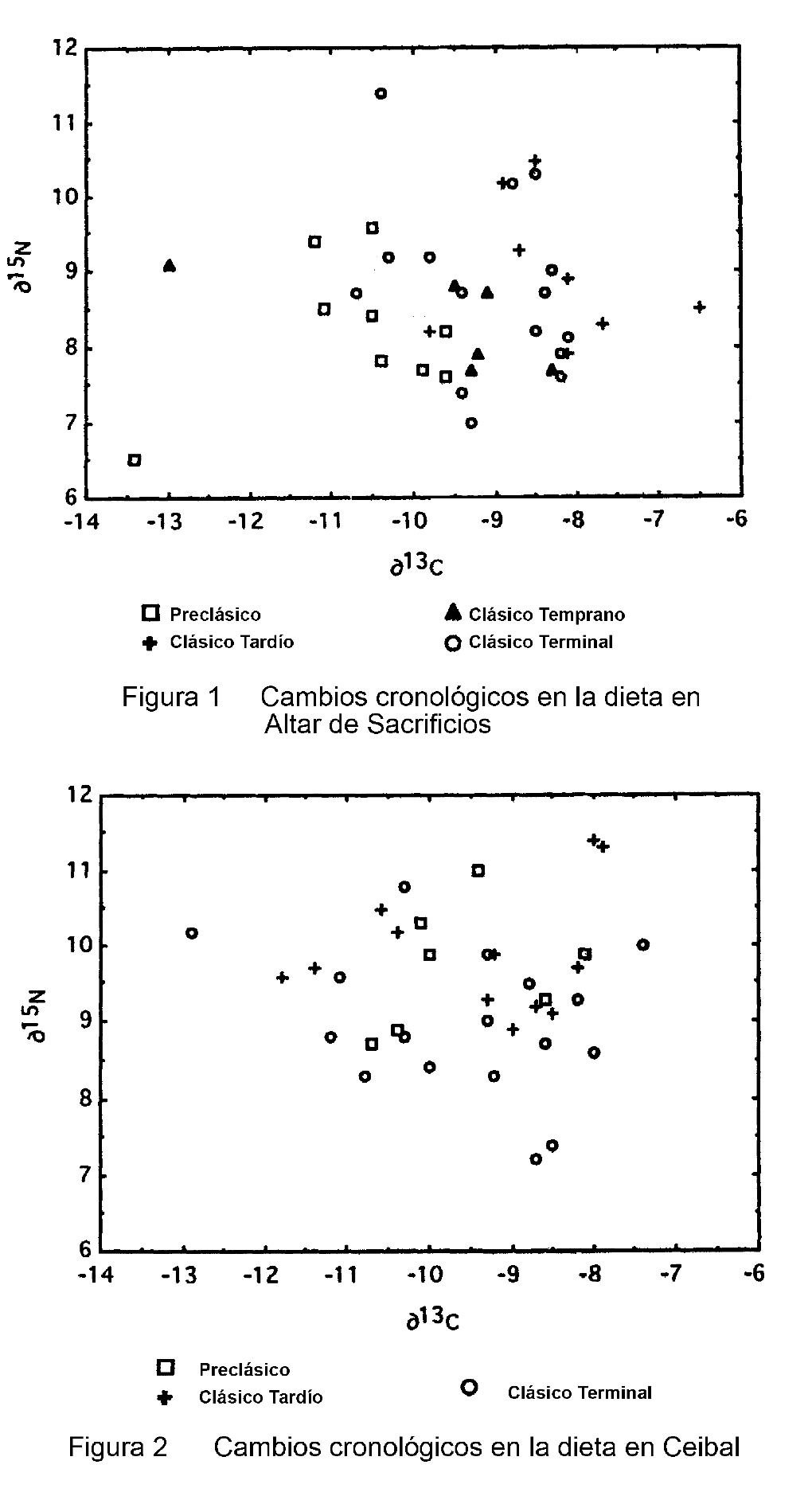
Los sitios de Altar de Sacrificios y Ceibal fueron ocupados desde el periodo Preclásico hasta el Clásico Terminal y dan una oportunidad de investigar patrones cronológicos en la dieta. En Altar de Sacrificios hay una tendencia clara en δ13C a través del tiempo (Figura 1). El nivel de carbono 13 es similar en esqueletos del Periodo Preclásico y Clásico Temprano (F=1.74, p=0.21) pero aumentó en el Clásico Tardío (F=4.27, p=0.06). En el Clásico Terminal, el nivel regresó cerca al nivel del Clásico Temprano, un cambio significativo en relación al nivel del Clásico Tardío (F=4.01, p=0.06). Aunque las últimas diferencias no son confirmadas al p0.05, claramente alcanzan significado. Este patrón indica que el consumo de plantas C4 (maíz) subió en importancia durante el periodo Clásico cuando el sitio estaba más poblado; pero cayó otra vez, en el Clásico Terminal, antes de la caída del sitio. Cambios en el nivel del nitrógeno 15 durante estas épocas eran nosignificativas, lo cual indica que este patrón refleja solamente cambios en el consumo relativo de maíz y otras plantas, no tanto en las fuentes de proteína explotadas.
En contraste, en Ceibal no hay un cambio obvio en δ13C entre periodos cronológicos, indicando que el consumo de maíz era estable a pesar de cambios significativos en la historia del sitio (Figura 2). Pero, la muestra de huesos del periodo Clásico Terminal se caracteriza por un nivel ligero de δ15N con respecto al nivel del Clásico Tardío (F=7.184, p=0.01). En comparación a tiempos anteriores, este resultado sugiere un mayor consumo de plantas leguminosas o proteína terrestre, con menor consumo de proteína acuática, durante la última ocupación intensa del sitio.
En Dos Pilas, es probable que cuatro entierros del Grupo L43 provengan de una ocupación residual del Clásico Terminal, debido a la presencia de cerámica Naranja Fina en el relleno de la Estructura L450 y sobre la plataforma L471 donde se colocaron los entierros (Antonia Foias, comunicación personal 1992). Estos esqueletos son isotópicamente indistinguibles de los del Clásico Tardío en el sitio (δ13C: F=1.036, p=0.34; δ15N: F=0.32, p=0.59), un resultado que podría indicar la continuidad de prácticas dietéticas a pesar del colapso del sitio. Por otra parte, es posible que la ocupación terminal fuera breve y que cambios dietéticos durante esta época no pueden ser identificados contra los 10 años de historia dietética que está registrada en el hueso (Chisholm 1989). Desafortunadamente, el pequeño tamaño de la muestra limita la interpretación de estos datos.
Además de patrones diacrónicos, los datos isotópicos revelan diferencias sincrónicas entre los sitios. Durante el periodo Preclásico, entierros de Altar y Ceibal presentaron diferencias isotópicas en δ15N, (F=11.54, p=0.004), demostrando un mayor consumo de proteína acuática en la muestra de Ceibal que es sugerido por el δ15N pesado de esa muestra. El nivel de δ13C en los esqueletos de Ceibal es más pesado que en Altar de Sacrificios también, una diferencia
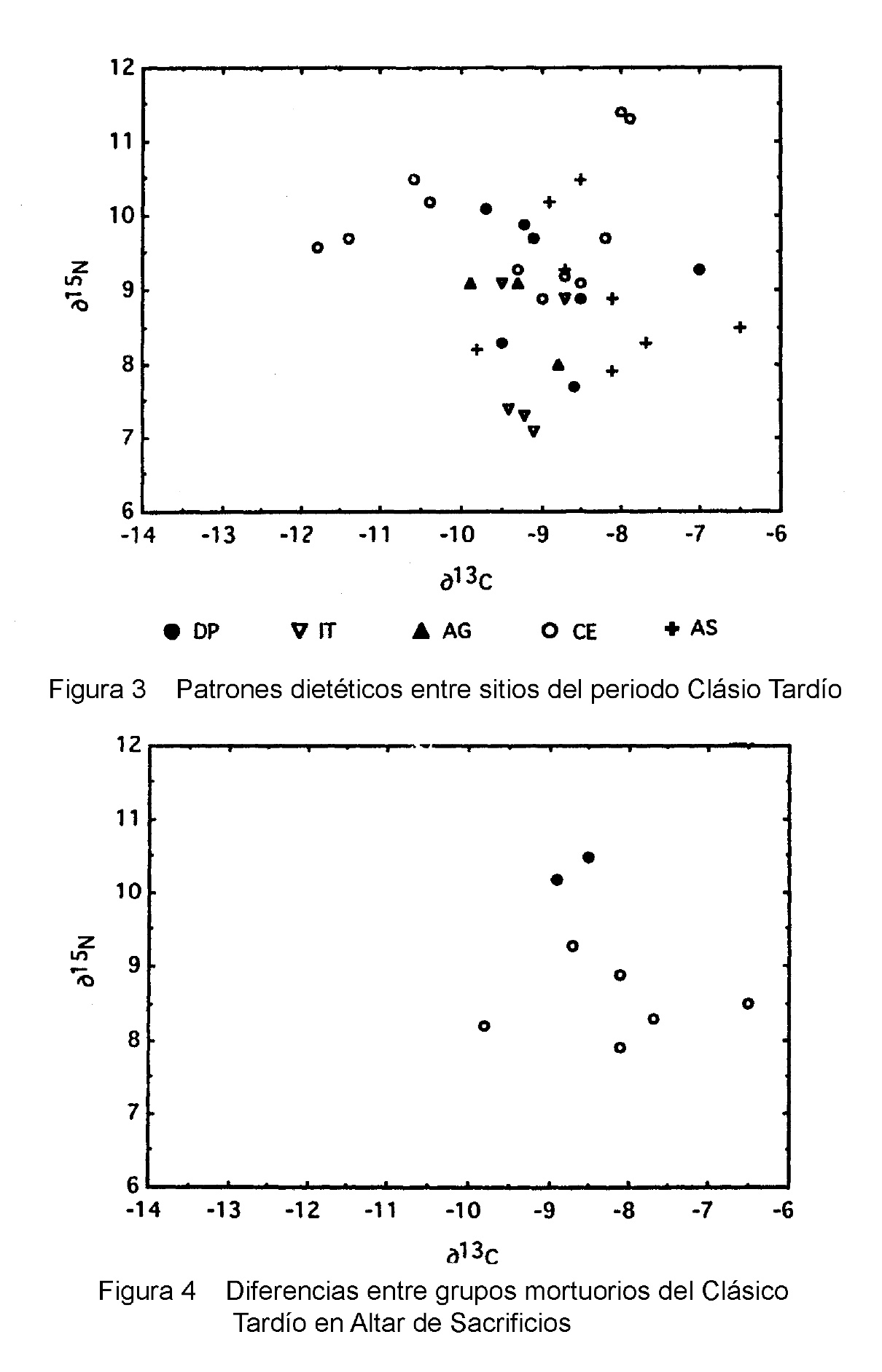
El Clásico Tardío presenta pocas diferencias dietéticas entre los sitios (Figura 3). Esqueletos de Ceibal son más pesados en δ15N en comparación con los demás sitios (F=12.15, p=0.001). Esta diferencia sugiere un consumo acuático mayor, una interpretación que es apoyada por el hecho de que los esqueletos de Ceibal pueden también ser ligeramente livianos en δ13C en comparación con otros sitios, aunque esta diferencia no es significativa estadísticamente (F=3.03, p=0.09). Diferencias entre los sitios restantes son pequeñas, e insignificantes para la comparación entre Dos Pilas y Altar de Sacrificios (F=0.11, p=0.74), o no pueden se probadas debido al tamaño pequeño de la muestra, como en el caso de Aguateca e Itzan.
En cambio, durante el Clásico Terminal, no aparecen diferencias significativas en carbono (F=0.68, p=0.51), ni nitrógeno (F=0.17, p=0.84), entre las muestras analizadas de Ceibal, Altar de Sacrificios y la pequeña muestra de Dos Pilas. Este resultado es muy interesante, en vista de las diferencias marcadas en la escala y organización política de las ocupaciones terminales de los sitios.
Además de las comparaciones entre los sitios, datos isotópicos puedan ser usados para examinar la distribución diferencial de alimentos dentro de la comunidad. Desafortunadamente, las dos muestras Preclásicas son muy pequeñas para investigar patrones sociales de consumo, tampoco éstas pueden ser agrupadas porque muestran diferencias entre ellas. Dentro de los sitios Clásicos Tardíos, excluyendo Ceibal, los esqueletos de hombres exhibieron niveles más pesados significativos de δ13C que en mujeres (F=5.05, p=0.04), pero niveles de δ15N son comparables (F=0.78, p=0.39). Diferencias entre los sexos no pueden ser examinados en la muestra de Ceibal porque solamente contiene una sola muestra de mujer. Este diferencia sexual en el consumo se mantiene en los muestras del Clásico Terminal (δ13C: F=10.25, p=0.003; δ15N: F=0.00, p=1.00). Estos resultados indican que los hombres consumieron proporcionalmente más maíz que las mujeres en la prehistoria tardía de la región del río Pasión, pero el acceso a la proteína fue disfrutado igualmente.
Acceso diferencial a los alimentos por miembros de diferente status está claramente ilustrado entre los enterramientos Clásicos Tardíos de Altar de Sacrificios (Figura 4). Los esqueletos de los Entierros 88 y 128, que fueron enterrados en criptas elaboradas y acompañados con más de 15 vasijas de cerámica, jade y cuentas de concha, son isotópicamente distintos a los entierros menos elaborados del Clásico Tardío. Ambos muestran un elevado δ15N sugiriendo un consumo mayor de proteína de agua dulce que aquel visto en otros esqueletos. Aunque el consumo de fauna marina pudo también haber producido este resultado (White y Schwarcz 1989), es improbable que alimentos marinos fueran conseguidos en esta área que está tan lejos de la costa.
DISCUSIÓN
Aunque es tentadora la interpretación de los datos mencionados en término de cambios de estrategias dietéticas en tiempo y espacio, algunos de los patrones pueden derivar de las diferencias de status social entre las muestras. Las series mortuorias investigadas aquí son el resultado de diferentes tipos de muestreo arqueológico. Si hubo una marcada variación en la dieta entre los miembros de una sola comunidad en un tiempo, como vemos entre los grupos mortuorios en Altar de Sacrificios y entre los sexos, la interpretación de datos comparativos entre sitios y épocas se vuelve más complicada. Se debe tener en cuenta las diferencias en la composición mortuoria entre las muestras, antes de comparar la dieta de entierros entre sitios y periodos cronológicos.
La demostración de diferencias de status en el consumo de alimentos en el Clásico Tardío de Altar de Sacrificios, paralela la identificación de acceso diferencial a recursos marinos sugerida por White y Schwartz (1989) en un esqueleto masculino del Clásico Temprano enterrado en una tumba en Lamanai, como también estudios isotópicos en la prehistoria norteamericana (Bender et al 1981; Schurr 1992). Por esta razón, la continuación del estudio de los datos de la región del río Pasión será enfocada hacia la identificación de grupos de status entre los restos mortuorios, con la finalidad de controlar esta fuente de variación.
A pesar de estas precauciones, es evidente que la dieta era afectada por cambios en la historia cultural de la civilización. Cambios en la dieta se documentaron en relación al desarrollo y colapso de Altar de Sacrificios y Ceibal, lo que sugiere que la dieta podría haber estado involucrada en la trayectoria de los sitios. Es interesante anotar que el colapso de la autoridad política ocurrió más tarde en Ceibal que en el resto de los sitios. Esta diferencia es también vista en el patrón dietético entre los sitios. El aumento gradual en el consumo de maíz en Altar de Sacrificios provee un fuerte contraste con el sitio de Lamanai, en Belice. Allí, el consumo de maíz declinó durante el periodo Clásico y el sitio es uno de los pocos que sobrevivió el colapso, siendo ocupado hasta tiempos históricos (White y Schwarcz 1989).
Aunque los resultados presentados aquí son preliminares, éstos dan alguna idea de la naturaleza de la dieta antigua de los Mayas. El maíz era la comida corriente durante todos los periodos de la ocupación Maya, pero esta era aumentada con una variedad de otros alimentos. Una caracterización mejor de los componentes de la dieta Maya será facilitada por el análisis isotópico de una muestra más grande de plantas y animales modernos actualmente en proceso. Además, las mismas fuentes de comida y entierros arqueológicos están siendo estudiadas dentro de un segundo tipo de análisis químico, la medición de abundancias elementales, que podría ofrecer mayor información de los patrones dietéticos. En particular, la relativa abundancia de estroncio y bario puede ser útil para separar la contribución de proteína terrestre y acuática a la dieta. En la medida que estos estudios progresen, se espera que ellos revelen una visión más completa de las implicaciones de la cocina Maya.
Este estudio fue hecho con el generoso permiso del Instituto de Antropología e Historia de Guatemala y el Museo Peabody de la Universidad de Harvard. También se agradece al Dr. A. Demarest, de la Universidad de Vanderbilt, por la oportunidad de trabajar junto con el Proyecto Arqueológico Regional Petexbatun. Agradecemos también a Kevin Johnston, de la Universidad de Yale, por su propuesta de examinar los entierros de Itzan. La ayuda de la Dra. M. Schoeninger, quien hizo el análisis isotópico, fue muy valiosa. La traducción de este artículo fue hecha con ayuda de M.C. Lozada C. Los análisis químicos se realizaron con el apoyo financiero de la Fundación Nacional de Ciencias (Estados Unidos) y la Fundación WennerGren. Durante el periodo de investigación, la autora ha recibido apoyo del Concejo de Investigaciones en Ciencias Naturales e Ingeniería (NSERC, Canadá), la Universidad de Chicago y la Fundación Canadiense de Mujeres Universitarias.
REFERENCIAS
Adams, Richard E. W.
1983 Ancient Land Use and Culture History in the Pasion River Region. En Prehistoric Settlement Patterns (editado por E. Z. Vogt y R. M. Leventhal):319335. University of New Mexico Press, Albuquerque.
Chisholm, B. S.
1989 Variation in Diet Reconstructions Based on Stable Carbon Isotopic Evidence. En The Chemistry of Prehistoric Human Bone (editado por T. D. Price):1037. Cambridge University Press, Cambridge.
Demarest, Arthur A.
1990 Warfare, Demography, and Tropical Ecology: Speculations on the Parameters of the Maya Collapse. Ponencia, American Anthropological Association, New Orleans.
Inomata, T., C. Beekman, S. Symonds, O. Chinchilla, F. Moscoso, L. Wright y R. Chatham.
1991 Excavations and Survey of the Petexbatun Fortress Capital of Aguateca. Ponencia, International Congress of Americanists, New Orleans.
Katzenberg, M. A.
1989 Stable Isotope Analysis of Archaeological Faunal Remains from Southern Ontario. Journal of Archaeological Science 16:319329.
Lentz, David L.
1991 Maya Diets of the Rich and Poor: Paleoethnobotanical Evidence from Copan. Latin American Antiquity 2 (3):269287.
Mathews, Peter y Gordon R. Willey
1991 Prehistoric Polities of the Pasión Region: Hieroglyphic Texts and their Archaeological Settings. En Classic Maya Political History: Hieroglyphic and Archaeological Evidence (editado por T. Patrick Culbert):30-71. School of American Research Advanced Seminar Series. Cambridge University Press, Cambridge.
DeNiro, M. J.
1985 Postmortem Preservation and Alteration of in Vivo Bone Collagen Isotope Ratios in Relation to Palaeodietary Reconstruction. Nature 317:806809.
1987 Stable Isotopy and Archaeology. American Scientist 75:182191.
Chisholm, B. S.
1989 Variation in Diet Reconstructions Based on Stable Carbon Isotopic Evidence. En The Chemistry of Prehistoric Human Bone (editado por T. D. Price):1037. Cambridge University Press, Cambridge.
Pohl, Mary
1985 The Privileges of Maya Elites: Prehistoric Vertebrate Fauna from Seibal. En Prehistoric Lowland Maya Environment and Subsistence Economy (editado por Mary Pohl):133145. Papers of the Peabody Museum of Archaeology and Ethnology, Vol. 77. Harvard University Press, Cambridge.
Price, T. D., M. Connor y J. D. Parsen
1985 Bone Chemistry and the Reconstruction of Diet: Strontium Discrimination in WhiteTailed Deer. Journal of Archaeological Science 12:419442.
Puleston, Dennis E.
1982 The Role of Ramon in Maya Subsistence. En Maya Subsistence (editado por K. Flannery):353367. Academic Press, New York.
Schurr, M. R.
1992 Isotopic and Mortuary Variability in a Middle Mississippian Population. American Antiquity 57 (2):300320.
Schwarcz, H. P. y M. J. Schoeninger.
1991 Stable Isotope Analyses in Human Nutritional Ecology. Yearbook of Physical Anthropology 34:283321.
Sharer, Robert J.
1978 The Prehistory of Chalchuapa, El Salvador. University of Pennsylvania Press, Philadelphia.
Van der Merwe, N. J.
1982 Carbon Isotopes, Photosynthesis, and Archaeology. American Scientist 70 (6):596606.
White, C. D. y H. P. Schwarcz
1989 Ancient Maya Diet: As Inferred from Isotopic and Chemical Analysis of Human Bone. Journal of Archaeological Science 16:451474.
Willey, Gordon R.
1973 The Altar de Sacrificios Excavations, General Summary and Conclusions. Papers of the Peabody Museum of Archaeology and Ethnology, Vol. 64, No. 3. Harvard University, Cambridge.